Reflexbogen (Physiologie)
Als Reflexbogen wird in der Physiologie die kürzeste Verbindung zwischen Rezeptor und Effektor über die Nervenzellen eines bestimmten neuronalen Erregungskreises bezeichnet.[1] Die Verschaltung vom afferenten auf das efferente Neuron erfolgt im einfachsten Fall auf spinaler Ebene über eine Synapse im Vorderhorn des Rückenmarks. Man spricht daher bei dieser Form eines Reflexes auch genauer von einem einfachen monosynaptischen Reflexbogen.[2]
Propriozeption und Regelkreis

Die Bezeichnung Reflexbogen und die damit verbundene physiologische Neuronentheorie ist angelehnt an das Konzept des technischen Regelkreises und an entsprechende Input/Output-Systeme. Im Gegensatz zum rein physiologischen Begriff des Reflexes wird mit dem Begriff des Reflexbogens das biologische Organisationsprinzip in topologischer Hinsicht betont. Die genaue Kenntnis topographischer Gegebenheiten ermöglicht eine exakte topische Diagnostik. Sie wird dadurch erleichtert, dass der gesamte monosynaptische Reflexbogen im gleichen Rückenmarksniveau liegt, dessen genaue Kenntnis demnach auch klinisch wichtig ist. Die Prüfung der Reflexe gehört zur neurologischen Standarduntersuchung.
Die Problematik einer topischen Diagnostik und der damit verbundenen Lokalisationslehre klingt auch mit dem Begriff der Proprioception bzw. mit dem Prinzip der Selbststeuerung oder Selbstregulation an.[3][4] Auch wenn nur ein kleiner Teil der Impulse der Propriozeption zum Bewusstsein, und damit zur Hirnrinde gelangt, so darf das Konzept des Reflexbogens nicht ausschließlich im Sinne eines simplen mechanischen Automatismus verstanden werden. Dies wäre eine unzulässige Vereinfachung, die dem Wesen des lebenden Organismus nicht gerecht wird. Eine einzelne Nervenzelle empfängt nicht nur Erregungen von ein oder zwei Neuronen, sondern von zahlreichen, ja bis zu Tausenden von Neuronen. Dies gilt auch für die motorische Vorderhornzelle im Rückenmark, vgl. den folgenden Abschnitt Elemente des Reflexbogens. So erhält z. B. der Reflexbogen fördernde oder hemmende zentrale Einflüsse durch die Pyramidenbahnen, die sich am peripheren motorischen Neuron in den Reflexbogen einschalten. Sie wirken physiologischerweise beim Eigenreflex hemmend auf eine Reflexantwort, bei Schädigung des Pyramidenbahnsystems dagegen fördernd, vgl. Pyramidenbahnzeichen.
Von der Vielzahl der tatsächlichen Reflexbögen sei hier hauptsächlich der monosynaptische Eigenreflex dargestellt.
Elemente des Reflexbogens

Entsprechend genanntem Prinzip eines Regelkreises wird unterschieden:
- auf der Seite der Eingabe (Bild „Regelkreis“, Symbol w) der afferente Schenkel des Bogens (blau in Bild „Querschnitt“); Ursprung = Sensor oder Fühler in der Technik = Rezeptor (Muskelspindel) in der Biologie; Weiterleitung des Reizes durch unipolare Nervenzelle im Spinalganglion
- auf der Seite der Ausgabe (Bild „Regelkreis“, Symbol y) der efferente Schenkel des Bogens (rot in Bild „Querschnitt“); Weiterleitung der Reizantwort durch motorische Vorderhornzelle; Ziel = Aktor bzw. Effektor in der Technik = Effektor (Physiologie) oder Wirkorgan in der Physiologie (Muskel oder Drüse)
Afferenzen haben bei einfachen (monosynaptischen) Reflexen ihren Ursprung in Sinnesorganen oder sonstigen sensiblen oder sensorischen Rezeptoren in Muskeln (Rezeptor: Muskelspindeln), Sehnen oder in der Haut (Tastsinn). Die Weiterleitung der afferenten Impulse zum Rückenmark erfolgt über sensible Nervenzellen (meist Aα-Fasern nach Erlanger Gasser bzw. Ia-Fasern/Ib und II-Fasern nach Lloyd/Hunt). Es handelt sich hinsichtlich des neuronalen Zelltyps um pseudounipolare Nervenzellen, deren Zellkörper im Spinalganglion (Bild „Querschnitt“, Ziffer 13: Spinalganglion) liegt. Dieses befindet sich innerhalb des Spinalkanals, gehört aber nicht zum zentralen, sondern zum peripheren Nervensystem, siehe die Definition des polysynaptischen Reflexes.
Efferenzen haben ihr Ziel in Muskel oder Drüse. Die Weiterleitung der efferenten Impulse vom Rückenmark erfolgt über motorische Nerven (Motoneuronen), deren Zellkörper im Bereich der grauen Substanz des Rückenmarks im motorischen Vorderhorn (Bild „Querschnitt“, Ziffer 1: Vorderhorn) liegt. Das zum Effektor (Muskel) führende Motoaxon gehört hinsichtlich der Leitgeschwindigkeit zu den Aα-Fasern (kurz: α-Motoneuron). Die Muskelspindeln werden motorisch über verschiedene Typen von Aγ-Fasern versorgt (kurz: γ-Motoneurone).
Physiologische Komponenten
Die Reflexantwort eines Lebewesens im Sinne des Reflexbogens besteht insgesamt aus folgenden funktionellen Einzeltatsachen:
- Registrierung eines adäquaten (meist als „auslösend“ bezeichneten, mechanisch evtl. auch so gedeuteten) Reizes oder Stimulus, auf den dieses Lebewesen von sich aus reagiert,
- Weiterleitung der dadurch zustande kommenden neuronalen Aktivierung oder „Erregung“ der jeweiligen sensorischen Nerven zu ihrem spezifischen Verarbeitungszentrum (Reflexzentrum) im Strickleiternervensystem bei niederen bzw. im Rückenmark bei höher entwickelten Tieren,
- Synaptische Überleitung der einlaufenden Erregung auf anatomisch festliegende, mit den Afferenzen verbundene motorische Nerven, über welche die jeweilige Reflexantwort zustande kommt oder „in Gang gesetzt wird“,
- Weiterleitung der Aktivierung der betreffenden motorischen Nerven zu den Muskeln oder Drüsen (als Effektoren), deren Zusammenwirken zu der reflektorischen Antwort (engl. response) führt und
- Aktivierung dieser Effektoren, wodurch der Reflex (bei Muskeln als Effektor bzw. als eine reflexhafte körperliche Bewegung) zustande kommt, die den jeweiligen Reflex definiert.
Weitere Begriffe
Gelegentlich spricht man auch von einem Reflexkreis, wobei allerdings auch reflektorische Zusammenhänge komplexerer Art und durch ineinander verschachtelte Regelkreise gemeint sein können. Die Zeit vom Einwirken des Reizes bis zur motorischen Antwort wird als Reflexzeit bezeichnet.
Mono- und polysynaptische Reflexe
Monosynaptische Reflexe
Über monosynaptische Reflexbögen (Eigenreflexe) zustande kommende Reflexreaktionen sind die am schnellsten ablaufenden organismischen Reflexe, da sie nur über eine einzige Synapse im Rückenmark laufen. Als Rückenmarksreflexe können sie in ihrem Verlauf nicht willentlich beeinflusst werden, wenn sie einmal in Gang gekommen sind; eine bewusste Beeinflussung gelingt nur indirekt über eine Reizkontrolle. Diese monosynaptischen Reflexe werden auch Eigenreflexe genannt, weil Rezeptor und Effektor im selben Organ liegen.
Beispiele für Eigenreflexe:
- Patellarsehnenreflex = PSR
- Achillessehnenreflex = ASR
- Kritische Anmerkung: Die Bezeichnung „Sehnenreflex“ ist pragmatisch, weil sie die Art und Weise der Auslösung beschreibt, nämlich durch einen Schlag auf die Sehne mit Hilfe des Reflexhammers. Diese Namensgebung erscheint jedoch leider unphysiologisch, da „Sehnenreflexe“ immer „Muskeldehnungsreflexe“ sind. Daher werden häufig anstelle der praktischen Bezeichnungen PSR und ASR u. a. die physiologisch korrekten Bezeichnungen Quadrizepsreflex für PSR und Triceps-surae-Reflex für ASR vorgezogen.[1] Weitere kritische Anmerkungen zur klassischen pragmatisch-experimentellen Reflextheorie siehe Kap. Reflexbogen als Modell.
Polysynaptische Reflexe
Bei den polysynaptischen Reflexen sind mehrere zentrale Neurone hintereinander geschaltet. Rezeptor und Effektor liegen meist räumlich getrennt, so dass sie auch als Fremdreflexe bezeichnet werden.[5]
Beispiele für Fremdreflexe:
- Beim Husten wird der Reiz durch Fremdkörper im Hals (Rezeptor) aufgenommen. Die Reizantwort erfolgt durch die Atemmuskulatur (Effektor).
- Beim Cornealreflex wird die Hornhaut des Auges (Rezeptor) berührt, die Reizantwort erfolgt durch die Lidmuskulatur (Effektor).
Reflexbogen als Modell
Der Reflexbogen als physiologisches Modell ist für das Verständnis der Arbeitsweise des gesamten Nervensystems von weitreichender Bedeutung. Dies trifft bereits zu auf das (im Vorspann) definitionsgemäß gegebene begriffliche Verständnis des Reflexbogens als Sonderfall der weit komplexeren Arbeitsweise neuronaler Erregungskreise. Das vegetative und das animalische Nervensystem sind jeweils nach diesem Prinzip tätig, stellen aber als solche Untereinheiten nur Elemente innerhalb des gesamten Nervensystems dar. Anhand der Funktion neuronaler Netzwerke können die Tätigkeiten der höheren Zentren des ZNS grundlegender verstanden werden, so z. B. auch die Unterscheidung zwischen Assoziationscortex und Sensomotorischem Cortex. Projektionszentren können z. B. in Analogie zu den Rückenmarkszentren als Teil eines „psychischen Reflexbogens“ (Jaspers) verstanden werden, siehe auch die Theorie der Spinalirritation von Wilhelm Griesinger.[6] Die Funktion des Assoziationscortex kann in Analogie zum polysynaptischen Reflex verstanden werden.[5][7]
Dieser Modellcharakter erstreckt sich somit auch auf das Verständnis psychischer Gegebenheiten einschließlich psychischer Krankheiten und Störungen. Letztere können z. T. als Versagen der Homöostase bzw. als Versagen unbewusster Selbstregulationen verstanden werden.[8][9] Bereits der moderne Begriff einer psychischen Störung weist solche Parallelen zu technischen Begriffen aus der Regeltechnik wie dem der Störgröße auf. Biologische Störgrößen entsprechen der Auseinandersetzung mit den wechselnden Situationen der Umwelt.[4] Auf diese Weise kann ebenso Aufschluss erhalten werden über körperliche Regelmechanismen bei psychischen Erkrankungen. Diese Sichtweise betrifft vor allem Fragen der psychophysischen Korrelation.
Die Beschreibung des bedingten Reflexes durch den russischen Wissenschaftler Iwan Petrowitsch Pawlow (1849–1936) stellt wissenschaftsgeschichtlich bereits einen ersten Schritt in diese Richtung dar. Hierbei wurden Phänomene der Wahrnehmung sowie des Lernens in die Theorie des Reflexes einbezogen. Am Beispiel Pawlows zeigen sich auch gesellschaftspolitische (materialistische) Hintergründe seiner Reflexlehre, die bisweilen auch von seinen Gegnern als „Reflexmythologie“ bezeichnet wurden.[4] Pawlow versuchte bei seiner Vorliebe für die Physiologie, alle psychologischen Termini aus seinem und dem Wortschatz seiner Mitarbeiter zu verbannen. Für seine Arbeiten über die Tätigkeit der Verdauungsdrüsen erhielt er 1904 den Nobelpreis.[10] Auch moderne Darstellungen dieses geschichtlichen Phänomens, das auch in der bundesdeutschen Vergangenheit für Bewegung sorgte, zeigen die unterschiedliche Ausdeutung kybernetischer Fragestellungen.[11] Die Einführung kybernetischer Begrifflichkeit kann zu einer Bereicherung der Beschreibung psychischer Störungen führen, die in der Sprache der klassischen Physik nicht möglich ist. Begriffe der Regeltechnik wie z. B. „In-Dienst-Nehmen“ verdeutlichen integrative Funktionen des ZNS (u. a. auch vergleichbar mit Faktoren sozialen Einflusses und entsprechender „In-Dienst-Nahme“) und den mit bestimmten technischen Lösungen vergleichbaren modularen Aufbau des Nervensystems. Auch in den Neurowissenschaften gibt es den Begriff der Module. Je komplexer diese Aufgaben sind, umso eher können sie als psychische Funktionen gewertet werden, was auch in der Praxis häufig dann passiert, wenn man sich bestimmte Reaktionen eines Menschen nicht erklären kann.[4][12]
- Funktionskreis-c.svg
Funktionskreis nach Jakob von Uexküll als Regelkreis auf der vegetativen Stufe
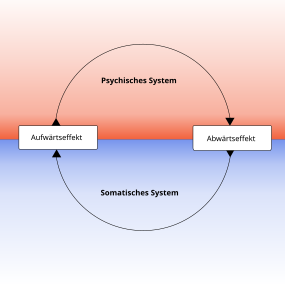
Psychophysische Korrelation als Regelkreis auf der animalischen Stufe

Die klassische Reflexlehre Pawlows, wie sie auch dem Behaviourismus zugrunde liegt, geht von der experimentellen Beobachtung im Labor aus, siehe Experimentelle Psychologie. John B. Watson (1878–1958) legte 1913 das Leitbild dieser neuen Wissenschaft fest, die sich später auch als Verhaltenswissenschaft oder als Verhaltensanalyse bewährt hat.[13] Dabei wurden zunächst die äußerlich zu beobachtenden physiologischen Abläufe untersucht, siehe blauer Bereich in Bild „Funktionskreis“ („Außenwelt“). Die auf das biologische System („roter Bereich“) afferent bzw. rezeptorisch von außen einwirkenden physikalischen Einflüsse wurden als Reiz (engl. stimulus) benannt, die von dem biologischen System ausgehenden efferenten bzw. effektorischen Reizantworten als Reaktionen (engl. response oder reaction). Dabei kam dem Reiz eine bestimmende, gleichsam kausale und vordergründige Bedeutung zu. Diese vorwiegend durch das Maschinenparadigma bestimmten Vorstellungen bedurften jedoch einer Revision, da auch dem biologischen System eine eigene Wirksamkeit und Rolle bei diesen Vorgängen zukommt. Die sogenannten S/R-Verknüpfung (engl. stimulus/response-model) bei biologischen Organismen ist demnach von rein physikalischen Vorgängen nach dem Ursache-Wirkungsmechanismus zu unterscheiden, wenngleich sie eine solche Deutung nahelegt. Bei den eigengesetzlichen (sogenannte „primär aktiven“) Abläufen biologischer Systeme sind auch die polysynaptischen Verbindungen bei Reflexabläufen zu berücksichtigen, insbesondere die Einflüsse des vegetativen und animalischen Nervensystems und ihrer Reaktionsbereitschaft.[14] Zusätzlich zu den angeborenen Reflexabläufen müssen nicht nur erlernte Einflüsse beachtet werden (Konditionierung), auch die allgemeine Entwicklung und Reifung des untersuchten biologischen Systems spielt dabei eine Rolle, siehe den blauen Bereich in Bild „Funktionskreis“ („Innenwelt“ – vegetatives NS) und die in Bild „Psychophysische Korrelation“ („psychisches System“ – animalisches NS) weiter aufgeteilte Gliederung dieser Innenwelt in einen somatischen und psychischen Bereich. Die „Innenwelt“ des Organismus in Bild „Funktionskreis“ entspricht sowohl dem „somatischen System“ als auch dem „psychischen System“ in Bild „Psychophysische Korrelation“ vgl. auch die nur vorläufige Hilfskonstruktion der Black-Box. Diese „Kiste“ wurde deshalb „schwarze Kiste“ benannt, weil die Faktoren des Innenlebens aus methodischen Gründen ausgeklammert werden sollten. Als Beispiel für die Wirksamkeit der Innenweltfaktoren (dem negativen Inbegriff der „Black Box“) wäre hier der Babinski-Reflex zu nennen. Dieser ist in seinem Auftreten an die Funktion zentraler ausgereifter Bahnen gebunden. Beim ausgereiften Erwachsenen fehlt er normalerweise, beim Kind ist er bis zum 2. Lebensjahr physiologisch.[1][15][16]
Eine frühe Kritik der behavioristischen Reflexbogentheorie findet sich bei dem Neurologen und Gestalttheoretiker Kurt Goldstein. Durch seine Arbeit mit hirngeschädigten Soldaten des Ersten Weltkriegs kommt er u. a. zu dem Ergebnis, dass es keine isolierten Reiz-Reaktions-Vorgänge im Organismus gibt, sondern dass der Organismus immer als Ganzes reagiert.[17]
Kurt Goldstein beeinflusst mit seinen Arbeiten deutlich die Entwicklung der Gestalttherapie. Fritz Perls und Laura Perls, die Begründer der Gestalttherapie, beziehen sich direkt auf Goldstein.[18]
Der Modellcharakter des Reflexbogens und seine weitreichende Verdeutlichung anhand bionischer Beispiele lässt dennoch viele Fragen offen. Wenn auch bei alltäglichen Vergleichen etwa die Ähnlichkeit zwischen Zentralnervensystem und Telefonzentrale durch die Analogie zwischen polysynaptischem Reflexbogen und telefonischer Kommunikation erhellt wird, so ist doch etwa die Frage der Kommunikation bei Pflanzen ein unberücksichtigtes und gänzlich anderes Phänomen. Pflanzen verfügen bekanntlich über kein Nervensystem. Sie haben aber vielfach ähnliche biologische Aufgaben zu bewältigen wie auch Tiere und Menschen. Der Reflexbogen kann somit nur z. T. als übergreifendes Modell in der Biologie dienen. Für das Pflanzenreich etwa gelten hier ganz andere Bedingungen.[19]
Einzelnachweise
- ↑ a b c Friedrich Wilhelm Bronisch: Die Reflexe. Thieme, Stuttgart 1979; (a) zu „Reflexbogen Begriffsdefinition“, S. 3; (b) zu Stw. „Kritik an der Reflexbezeichnung allein nach der klinischen Methode ihrer Auslösung“ (Pragmatismuskritik), S. 5; (c) zu Stw. „Babinski-Reflex“ S. 71 f.
- ↑ einfacher monosynaptischer Reflexbogen. In: Peter Duus: Neurologisch-topische Diagnostik. Anatomie, Physiologie, Klinik. 5. Auflage. Georg Thieme Verlag, Stuttgart 1990, ISBN 3-13-535805-4, S. 11.
- ↑ Hans-Georg Gadamer: Über die Verborgenheit der Gesundheit. (Bibliothek Suhrkamp, Band 1135). Frankfurt am Main 1993; zu Stw. „Selbstbewegung“ und „heauto kinoun“ (Aristoteles): Kap. Leben und Seele, S. 179.
- ↑ a b c d Thure von Uexküll: Grundfragen der psychosomatischen Medizin. Rowohlt Taschenbuch, Reinbek bei Hamburg 1963; (a) zu Stw. „Selbstregulation“: Kap. Selbstregulation und Regelkreis, S. 251 ff.; (b) zu Stw. „Systematische Beispiele für biologische Störgrößen“, S. 262 f.; (c) zu Stw. „Reflexmythologie“, S. 165 ff.; (d) zu Stw. „Begriffsklärung des Psychischen und Komplexität gesteuerter Abläufe“ (Begriffliche Übertragung von „Psyche“ auf technisches Modell der Führungsgrößenaufschaltung), S. 261 f.
- ↑ a b Robert F. Schmidt (Hrsg.): Grundriß der Neurophysiologie. 3. Auflage. Springer, Berlin 1979, ISBN 3-540-07827-4; (a) zu den Stw. „Polysynaptischer Reflex“ und „Fremdreflex“ Seite 126, dgl. zu den Stw. „monosynaptischer Dehnungsreflex“ und „Eigenreflex“, S. 123; (b) zu Stw. „Assoziationskortex“, S. 282.
- ↑ Karl Jaspers: Allgemeine Psychopathologie. 9. Auflage. Springer, Berlin 1973, ISBN 3-540-03340-8; zu Stw. „psychischer Reflexbogen“: 1. Teil: Die Einzeltatbestände des Seelenlebens, 2. Kap.: Die objektiven Leistungen des Seelenlebens (Leistungspsychologie) b) Das neurologische Grundschema des Reflexbogens und das psychologische Grundschema von Aufgabe und Leistung, S. 130 ff.
- ↑ Manfred Spitzer: Geist im Netz. Modelle für Lernen, Denken und Handeln. Spektrum Akademischer Verlag, Heidelberg 1996, ISBN 3-8274-0109-7; zu Stw. „Mehrschichtige Netzwerke“: Kap. 6. Zwischenschichten, S. 125 ff.
- ↑ Homöostase. In: Stavros Mentzos: Neurotische Konfliktverarbeitung; Einführung in die psychoanalytische Neurosenlehre unter Berücksichtigung neuerer Perspektiven. Fischer Taschenbuch Verlag, Frankfurt 1992, ISBN 3-596-42239-6, S. 53, 187.
- ↑ Homöostase. In: Stavros Mentzos: Psychodynamische Modelle in der Psychiatrie. 2. Auflage. Vandenhoeck & Ruprecht, Göttingen 1992, ISBN 3-525-45727-8, S. 101.
- ↑ Pawlow, Iwan Petrowitsch. In: Wilhelm Karl Arnold u. a. (Hrsg.): Lexikon der Psychologie. Bechtermünz, Augsburg 1996, ISBN 3-86047-508-8, Spalte 1562 f.
- ↑ Kybernetik und Kulturgeschichte. In: Philipp Aumann: Mode und Methode. Die Kybernetik in der Bundesrepublik Deutschland. Wallstein-Verlag, Göttingen 2009, ISBN 978-3-8353-0449-9; Rezension
- ↑ Regeltechnik und Psyche. In: Karl Steinbuch: Automat und Mensch. Kybernetische Tatsachen und Hypothesen. 3. Auflage. Springer-Verlag, Berlin 1965, S. 7, 9, 12, 149, 281, 350, 395, 397, 401.
- ↑ John B. Watson: Psychology as the Behaviorist Views It. In: Psychological Review, 20, 1913, S. 158–177, psychclassics.yorku.ca – auch enthalten in: John B. Watson: Behaviorismus. Köln 1968 bzw. Frankfurt am Main 1976
- ↑ Ludwig von Bertalanffy: General System Theory. George Braziller, New York 1968.
- ↑ Peter R. Hofstätter (Hrsg.): Psychologie. Das Fischer Lexikon. Fischer-Taschenbuch, Frankfurt am Main 1972, ISBN 3-436-01159-2; zu Stw. „Bedingter Reflex“, S. 62 ff.; zu Stw. „Vorbild eines mechanischen Apparats“(Maschinenparadigma), S. 70 f.
- ↑ Thure von Uexküll u. a. (Hrsg.): Psychosomatische Medizin. 3. Auflage. Urban & Schwarzenberg, München 1986, ISBN 3-541-08843-5; zu Stw. „Revisionsbedürftigkeit der klassischen Reflextheorie“: Kap. 1.3.1 Die emergenten Eigenschaften der biologischen Systemebene, S. 10 f.
- ↑ K. Goldstein: Der Aufbau des Organismus. The Hague 1934, S. 45 ff.
- ↑ s. u. a. F. S. Perls: Das Ich, der Hunger und die Aggression. 1946, Stuttgart 1969, S. 59 ff.
- ↑ Volker Arzt: Kluge Pflanzen. Wie sie locken und lügen, sich warnen und wehren und Hilfe holen bei Gefahr. Riemann, 2009, ISBN 978-3-570-01026-6; Audiodatei der Buchbesprechung mit dem Autor vom 28. Februar 2010
