Beggiatoa
| Beggiatoa | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Illustration von Beggiatoa alba: | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Beggiatoa | ||||||||||
| Trevisan 1842[1] |
Beggiatoa ist eine Gattung von Gammaproteobakterien, die zur Ordnung Thiotrichales (alias Beggiatoales) gehören. Diese Bakteriengattung war eine der ersten, die der russische Botaniker Sergei Winogradski (englisch auch Winogradsky) entdeckte. Während seiner Forschungen im botanischen Labor von Anton de Bary im Jahr 1887 stellte er fest, dass Beggiatoa Schwefelwasserstoff (H2S) als Energiequelle oxidiert und dabei intrazelluläre Schwefeltröpfchen bildet, wobei Sauerstoff der terminale Elektronenakzeptor ist und CO2 als Kohlenstoffquelle dient. Winogradski bezeichnete diese Form des Stoffwechsels als „Inorgoxidation“ (Oxidation anorganischer Verbindungen), heute Chemolithotrophie genannt. Die Beggiatoa-Arten leben in schwefelreichen Umgebungen auf Böden, sowohl im Süßwasser als auch im Meer, etwa in den hydrothermalen Schloten der Tiefsee und in verschmutzten Meeresumgebungen. Diese Entdeckung war die erste Entdeckung einer Lithotrophie.[2][3] Diese farblosen Bakterien bilden fadenförmige Kolonien (Filamente, wie bei filamentösen Cyanobakterien auch Trichome genannt)[4] und kann in einem Biofilm angeordnet sein, der mit bloßem Auge sichtbar ist und aus sehr langen weißen Fadenmatten besteht; die weiße Farbe ist dabei auf den gespeicherten Schwefel zurückzuführen. Die Beggiatoa-Bakterien sind dabei manchmal mit anderen Schwefelbakterien vergesellschaftet, z. B. mit der Gattung Thiothrix Winogradsky 1888 aus derselben Familie Thiotrichaceae Garrity et al. 2005.[5]
Formell beschrieben wurden zwei Arten von Beggiatoa: die Typusart Beggiatoa alba und Beggiatoa leptomitoformis, wobei letztere erst 2017 veröffentlicht wurde.[6][7] Diese Arten von Beggiatoa haben Zellen mit einem Durchmesser von bis zu 200 µm und gehören damit zu den größten Prokaryoten der Erde.[5]
Genom
Wegen des Mangels an Reinkulturen war lange Zeit nur wenig über die Genetik von Beggiatoa bekannt. Beggiatoa alba hat einen GC-Gehalt zwischen 40 und 42,7 mol%. Es gibt zwei oder drei Plasmide; die Genomgröße Stammes Beggiatoa alba B18LD beträgt etwa 3 Mbp (Megabasenpaare).[5] In einer wichtigen Studie über das Genom von Beggiatoa wurden die Sequenzen von zwei Einzelfilamenten eines vakuolisierten Stammes analysiert. Die optische Kartierung zeigte, dass die Genomgröße etwa 7,4 Mbp beträgt, und die Sequenzanalysen zeigten Stoffwechselwege für die Schwefeloxidation, die Nitrat- und Sauerstoffatmung sowie die CO2-Fixierung nach, was die chemolithoautotrophe Physiologie von Beggiatoa bestätigt. Darüber hinaus deutet die vergleichende Genomik auf einen horizontalen Gentransfer (HGT) von Speicher-, Stoffwechsel- und Gleitfähigkeiten zwischen Beggiatoa und Cyanobakterien hin.[8]
Auch das Genom des Stammes Beggiatoa leptomitiformis D-402T (alias DSM 14946) wurde vollständig sequenziert. Die Größe beträgt 4.265.296 bp (Basenpaare), dazu kommt ein Plasmid von 6.185 bp; beide Teile zusammen kodieren nach Vorhersage 3.636 Gene.[9]
Taxonomie
Die Gattung Beggiatoa ist insgesamt eine ziemlich vielfältige Gruppe, deren Vertreter verschiedene Lebensräume und Nischen sowohl im Süß- als auch im Salzwasser besiedeln. In der Vergangenheit wurden sie aufgrund ihrer Morphologie und Motilität als Cyanobakterien der Gattung Oscillatoria klassifiziert,[10] aber Analysen der 5S (5S-RNA der ribosomalen RNA) von Stahl et al. (1987) haben gezeigt, dass die Mitglieder von Beggiatoa phylogenetisch weit von Cyanobakterien entfernt sind und zum Stamm der Gammaproteobakterien gehören.[11] Der erwähnte HGT mit Übernahme etlicher Cyanobakterien-Gene dürfte ein wesentlicher Grund für diese Übereinstimmungen und damit für die Unsicherheiten in der Taxonomie sein. Insbesondere ist die Zugehörigkeit weiterer ehemals als Mitglieder der vermeintlichen Cyanobakterien-Gattung Beggiatoa klassifizierten Spezies weiter in der Diskussion, denn diese können womöglich echte Cyanobakterien sein (s. u.).
Nach der
[6] und dem
[12] sind derzeit (Stand 6. Januar 2022) nur zwei Spezies der Gattung Beggiatoa gültig veröffentlicht worden: Beggiatoa alba und Beggiatoa leptomitoformis.
Die nächsten bekannten Verwandten von Beggiatoa sind Bakterien der Gattung Thioploca (Stand 1999).[13] Die beiden Gattungen Beggiatoa und Thioploca unterscheiden sich durch ein einziges morphologisches Merkmal: Die Thioploca-Filamente treten in Bündeln auf, die von einer gemeinsamen Hülle umgeben sind, während die Beggiatoa-Filamente als Einzelfilamente auftreten ohne diese Struktur auszubilden. Alle Beggiatoa- und Thioploca-Stämme sind in der Lage, Sulfid (bzw. H2S) zu elementarem Schwefel zu oxidieren, der in Form der intrazellulären Schwefelkügelchen gespeichert wird. Dadurch werden die Zellen sehr widerstandsfähig und unter dem Mikroskop auffällig. Diese Eigenschaft, zusammen mit dem Fehlen photosynthetischer Pigmente (Chlorophylle) unterscheidet die Gattungen Beggiatoa und Thioploca als fadenförmige Vertreter der „farblosen Schwefelbakterien“ von anderen fadenförmigen Bakterien, wie den Cyanobakterien und nicht-schwefeloxidierenden Heterotrophen wie Cytophaga und Flexibacter (beide Cytophagaceae, Abteilung Bacteroidetes). Entgegen früherer Auffassung gibt es keine enge evolutionäre Verwandtschaft zwischen Beggiatoa und Thioploca einerseits und anderen gleitenden Fadenbakterien andererseits.[5]
Ein weiteres charakteristisches Merkmal ist die Fähigkeit der weit verbreiteten marinen Arten von Beggiatoa und verwandter Bakterien, Nitrat in den Vakuolen der Zellen zu speichern. Untersuchungen der 16S rRNA-Sequenzen ergaben, dass dieses Merkmal von den Mitgliedern einer monophyletischen Gruppe innerhalb der Gattungen der Beggiatoa-Verwandtschaft geteilt wird. Zu dieser Gruppe gehören auch die Gattungen von Thioploca und Thiomargarita, die sich beide nur geringfügig von Beggiatoa unterscheiden: Während die Zellen von Thioploca in einer gemeinsamen Schleimhülle wachsen, hat Thiomargarita kein fadenförmiges Wachstum erhalten, sondern bildet Ketten aus runden Zellen. Da die überkommene Nomenklatur diese phylogenetische Geschichte nicht widerspiegelt, ist eine neue Benennung der Gattungen und Arten erforderlich.[13] Der neue Referenzstamm (englisch Neo-type strain) für die Süßwasserart Beggiatoa alba ist beispielsweise dann B18LB und dieser bestimmt die Kriterien für die Identifizierung dieser Spezies.[14]
Systematik
Äußere Systematik
Die Gattung Beggiatoa wird entweder als Mitglied der Familie Beggiatoaceae Migula 1894 (nach LPSN[6]) oder der (etwas größeren) Familie Thiotrichaceae Garrity et al. 2005 (nach NCBI[12] und WoRMS[15]) geführt.
Diese 2005 neu gefasste Familie Thiotrichaceae umfasst nach NCBI nicht nur die Beggiatoaceae, sondern auch die Familien Leucotrichaceae Buchanan 1957 und Achromatiaceae Massart 1901.
In beiden Fällen gilt die Beggiatoa enthaltende Familie als Mitglied der Ordnung Thiotrichales Garrity et al. 2005 Entweder gilt der Ordnungsname Beggiatoales Buchanan 1957 als Synonym von Thiotrichales (nach LPSN[6]), oder die Thiotrichales gelten als die Beggiatoales mit einschließend (nach NCBI[12]).
Mit Beggiatoa verwandte Gattungen in der Familie Beggiatoaceae bzw. Thiotrichaceae sind (Auswahl):
- Candidatus Allobeggiatoa Hinck et al. 2011 (LPSN: Beggiatoaceae, NCBI: Thiotrichaceae). Arten:
- Candidatus Allobeggiatoa salina Hinck et al. 2011 (LPSN)
- Candidatus Allobeggiatoa halophila Beutler et al. 2012 (LPSN, NCBI)
- Candidatus Halobeggiatoa Hinck 2009 (NCBI: Thiotrichaceae). Arten:
- Candidatus Halobeggiatoa sp. HMW-R907 (NCBI)
- Candidatus Halobeggiatoa sp. HMW-S1778 (NCBI)
- Candidatus Halobeggiatoa sp. HMW-S1880 (NCBI)
- Candidatus Halobeggiatoa sp. HMW-S2301 (NCBI)
- Candidatus Halobeggiatoa sp. HMW-S2528 (NCBI)
- Candidatus Halobeggiatoa sp. HMW-S2548 (NCBI)
- Candidatus Halobeggiatoa sp. HMW-S2829 (NCBI)
- Candidatus Halobeggiatoa sp. HMW-W520 (NCBI)
- Candidatus Halobeggiatoa sp. HMW-W562 (NCBI)
- Candidatus Halobeggiatoa sp. HMW-W572 (NCBI)
- Candidatus Isobeggiatoa Salman et al. 2011 (LPSN: Beggiatoaceae, NCBI: Thiotrichaceae). Arten:
- Candidatus Isobeggiatoa divolgata Salman et al. 2011 (LPSN, NCBI)
- Candidatus Isobeggiatoa sp. 'Guadeloupe FWI' (NCBI)
- Candidatus Maribeggiatoa Salman et al. 2011 (LPSN: Beggiatoaceae, NCBI: Thiotrichaceae). Art:
- Candidatus Maribeggiatoa vulgaris Salman et al. 2011 (LPSN, NCBI) – mit Stämmen Beggiatoa sp. 'Carmel Canyon' und Beggiatoa sp. 'Monterey Canyon' (NCBI)
- Candidatus Parabeggiatoa Salman et al. 2011 (LPSN: Beggiatoaceae, NCBI: Thiotrichaceae). Art:
- Candidatus Parabeggiatoa communis Salman et al. 2011 (LPSN, NCBI)
- Thiothrix nivea (Rabenhorst 1865) Winogradsky 1888, ehemals Beggiatoa nivea (LPSN)
- Thiomargarita Schulz et al. 1999
- Ca. Thiolava Danovaro et al. 2017 (LPSN)
Anm.: AlgaeBase klassifiziert die Gattung Beggiatoa unspezifisch in Taxa der Ränge Familie bis Reich (en.
Arten und Stämme
Quellen der folgenden Artenliste ist LPSN,[6] AlgaeBase,[16] WoRMS,[15] NCBI[12] (U=unsicher, P=provisorisch)
Gattung Beggiatoa V.B.A.Trevisan 1842/1845, mit Schreibvarianten Beggiatoaea, Beggiatonia
- Beggiatoa alba (Vaucher 1803) Trevisan 1845 (LPSN, AlgaeBase:U, WoRMS:U) – Typusspezies (LPSN) bzw. Lectotype (AlgaeBase), syn. Oscillatoria alba Vaucher 1803 inkl. Stamm Beggiatoa alba B18LD (NCBI)
- Beggiatoa leptomitiformis Dubinina et al. 2017 (LPSN) bzw. Trevisan 1842, mit Schreibvariante Beggiatoa leptomitoformis, inkl. Stamm Beggiatoa sp. D-402T alias DSM 14946[9][17] (NCBI)
- Beggiatoa minima Vinogradskij [Winogradsky] (AlgaeBase:U, WoRMS:U)
- Beggiatoa media Winogradsky (AlgaeBase:P, WoRMS:U)
- Beggiatoa punctata (Corda 1835) Trevisan 1842 (LPSN:P, AlgaeBase:P, WoRMS:U)
- Beggiatoa sp. 'Guadeloupe FWI'
- Beggiatoa sp. 'Guaymas Basin' mit Isolat Beggiatoa GUB8[19]
- Beggiatoa sp. 'Gulf of Mexico' mit Isolat Beggiatoa GOM9[20]
- Beggiatoa sp. 'Orange Guaymas'[21] (nach Winkel et al. 2016 zu Maribeggiatoa)[22]
- Beggiatoa sp. 1.1
- Beggiatoa sp. 11.1
- Beggiatoa sp. 13.1
- Beggiatoa sp. 1401-13
- Beggiatoa sp. 35Flor[23][24][22]
- Beggiatoa sp. 401
- Beggiatoa sp. 4572_84
- Beggiatoa sp. AA5A
- Beggiatoa sp. Arauama I[25][26]
- Beggiatoa sp. Arauama II[25][26]
- Beggiatoa sp. B15LD
- Beggiatoa sp. B2
- Beggiatoa sp. CSO1
- Beggiatoa sp. IS2
- Beggiatoa sp. LPN
- Beggiatoa sp. MS-81-1c (obligat autotroph, s. u.)[27][28]
- Beggiatoa sp. MS-81-6 (fakultativ autotroph, s. u.)[27][28]
- Beggiatoa sp. OCN075
- Beggiatoa sp. OCN076
- Beggiatoa sp. OCN077
- Beggiatoa sp. OH-75-2a
- Beggiatoa sp. PS
- Beggiatoa sp. SS
(Diskutierte) Neuzuordnungen
Auswahl einiger Neuzuordnungen (Stand 6. Januar 2022):[6][16][15][12]
- Innerhalb der Gammaproteobakterien, Fam. Thiotrichaceae:
- Zu den Cyanobakterien, Fam. Oscillatoriaceae:
- Beggiatoa arachnoidea (C.Agardh) Rabenhorst ⇒ Oscillatoria arachnoidea C.Agardh ex Gomont (AlgaeBase)
- Beggiatoa leptomitiformis Trevisan, 1842 ⇒ Oscillatoria leptomitiformis Meneghini ex Gomont, 1892 (AlgaeBase, WoRMS)
- Beggiatoa tigrina (Roemer) Rabenhorst ⇒ Oscillatoria tigrina Roemer ex Gomont (AlgaeBase)
- Beggiatoa versatilis (Kützing) Trevisan ⇒ Oscillatoria versatilis Kützing ex Gomont (AlgaeBase)
- Beggiatoa hinnulea Wolle ⇒ Hypheothrix hinnulea Forti (AlgaeBase)
- Beggiatoa mirabilis Cohn, 1865 ⇒ Havrella mirabilis G.Breton & P.Saulot, 1986 (AlgaeBase: incertae sedis, nach WoRMS aber zu Oscillatoriaceae)
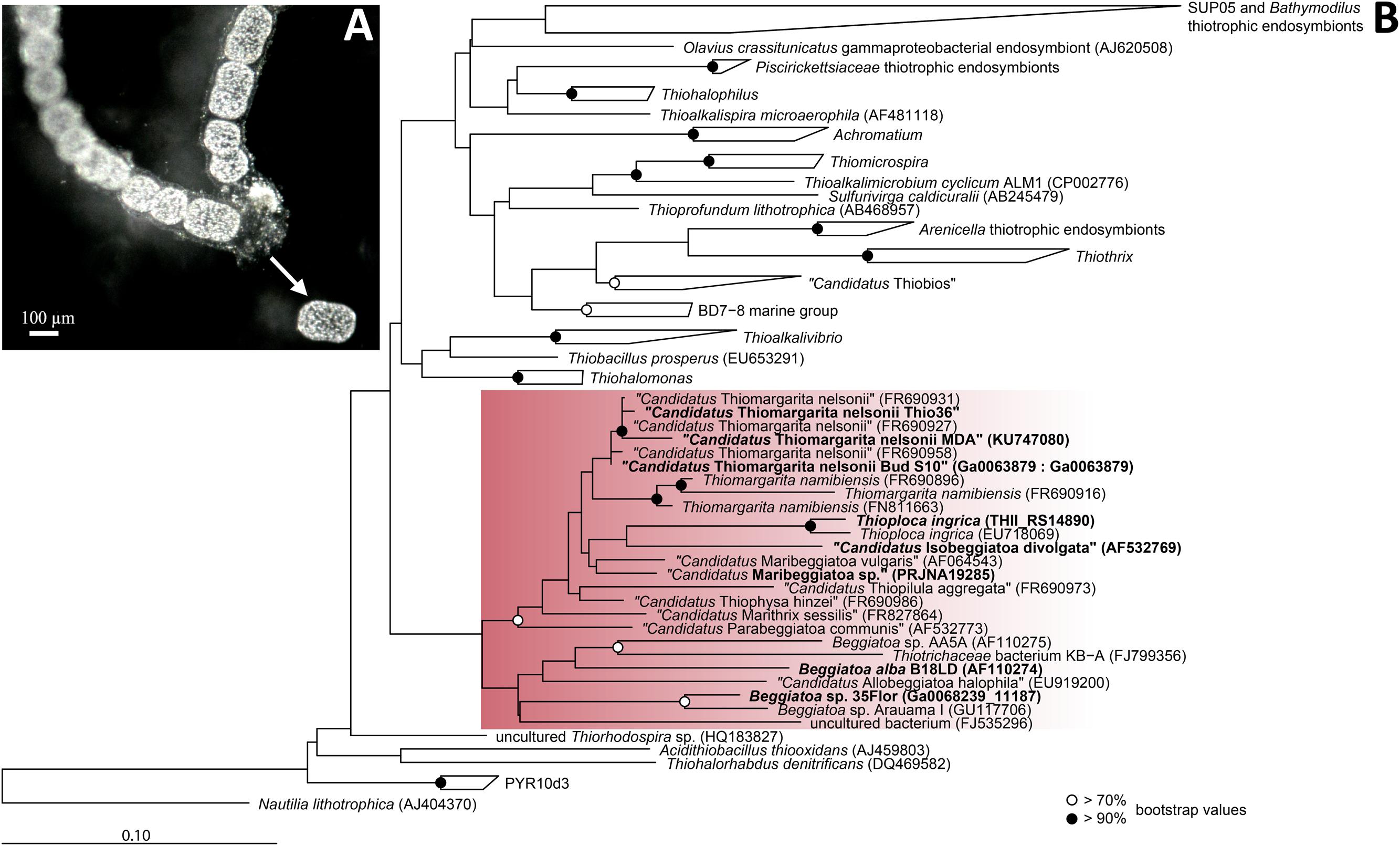
Über diese Neuzuordnungen hinaus erwies sich die Gattung Beggiatoa in den Jahren 2005 bis 2011 als nicht monophyletisch. Um das Problem zu beheben, wurden als neue Kandidatengattungen Isobeggiatoa, Maribeggiatoa, Parabeggiatoa und Allobeggiatoa vorgeschlagen, um einzelne bisherige Beggiatoa-Spezies aufzunehmen. Ein phylogenetischer Baum dieser und weiterer Spezies der Familie Thiotrichaceae findet sich bei der Gattung Thiomargarita §Äußere Systematik.
Etymologie
Winogradski benannte die Gattung zu Ehren von Francesco Secondo Beggiato,[2][3] einem italienischen Arzt und Botaniker aus Vicenza (1806-1883)[29], der die Abhandlung „
“ (Über die Euganeischen Thermen[30][31]) verfasste.[6]
Das Typusart-Epitheton kommt von lateinisch alba ‚die weiße‘.
Morphologie
Anhand ihrer morphologischen Merkmale lassen sich die Beggiatoa-Arten und -Stämme in drei Kategorien einteilen:
- Süßwasserstämme, die durch schmale Filamente ohne Vakuolen gekennzeichnet sind,
- schmale marine Stämme, ohne Vakuolen (Durchmesser der Fäden etwa 4,4 |µm),
- größere marine Stämme mit Vakuolen zur Speicherung von Nitrat (der Durchmesser der Filamente variiert zwischen 5 und 140 µm)
Natürlich ist diese Klassifizierung idealisiert, so dass es Ausnahmen geben kann. Schmale Filamente bestehen in der Regel aus länglich-zylindrischen Zellen, deren Länge etwa das 1,5- bis 8-fache ihrer Dicke beträgt, breitere Filamente bestehen hingegen aus eher scheibenförmige Zellen mit Längen, die das 0,10- bis 0,90-fache ihrer Zellbreite betragen. Bei allen kultivierten Stämmen sind die Endzellen der Filamente abgerundet. Obwohl es sich um gramnegative Bakterien handelt, weisen die Beggiatoa-Zellen eine ungewöhnliche Zellwand- und Membranorganisation auf. Manchmal gibt es weitere Membranen, die die Peptidoglykanschicht (Mureinschicht) bedecken, die Anzahl dieser Zusatzmembranen ist sehr unterschiedlich. Ihr Vorhandensein ist möglicherweise auf die rauen Bedingungen zurückzuführen, unter denen einige dieser Organismen leben. Sogar die intrazellulären Granula können von zusätzlichen Membran-Strukturen bedeckt sein. Neben den Schwefelkörnchen finden sich in den Zellen oft auch ähnlich gelagerte Polyhydroxybutyrat- und Polyphosphatkörnchen.
Sehr häufig in großen marinen vakuolierten (mit Vakuolen gefüllten) Beggiatoa sind „hohlstrukturierte“ Filamente (en.
), die aus schmalen Zellen mit einer großen Zentralen Vakuole zur Speicherung von Nitrat und wenig Zytoplasma bestehen.[5][27]
Motilität
Die Filamente bewegen sich gleitend, und diese Bewegung ist vermutlich mit fadenartigen Strukturen in der äußeren Membran und trans-Peptidoglykan-Kanälen verbunden. Manchmal können die Filamente auch (ähnlich wie bei den Cyanobakterien der Gattung Oscillatoria) durch die Bildung von Nekridien (Necridia, abgestorbenen Zellen, vgl. Nekrose) in der Mitte des Filaments durchbrechen. Die Motilität der Filamente ist für die Anpassungsfähigkeit der Bakterien von großer Bedeutung. Sie ermöglicht ihnen, sich in Richtung geeigneterer Bedingungen für den Zellstoffwechsel zu bewegen. Die wichtigsten Triebkräfte für die Bewegung der Beggiatoa-Filamente sind ein hoher Sauerstoff- und Sulfidgehalt (O2 und H2S), sowie Lichteinwirkung, von der sich die Filamente wegbewegen (negative Phototaxis).[27]
Zellwachstum und Vermehrung
Die Vermehrung von Beggiatoa geschieht durch Fragmentierung der Filamente (Trichome). Das Wachstum einer Kolonie führt zur Mattenbildung durch abwechselnde Fadenverlängerung und Fadenbruch.
Von grundlegender Bedeutung für die Fragmentierung sind (ähnlich wie bei den Cyanobakterien der Gattung Ocillatoria) nekrotische „Opferzellen“ (en.
), genannt Nekridien (oder Necridia), die durch ihr Absterben den Bruch ermöglichen.
Sie unterbrechen dadurch die Kommunikation zwischen den beiden Teilen eines Filaments unterbrechen, so dass jeder Abschnitt seine Gleitrichtung ändern kann, was in der Folge zum Auseinanderbrechen führt.[32][4]
Die durchschnittliche Filamentlänge, die durch diesen Prozess erreicht wird, ist auch das Ergebnis von Wechselwirkungen zwischen Genen und Umwelt. Beispielsweise hängen das Wachstum und Position des Filaments von vertikalen Gradienten von Sauerstoff und Sulfid ab. Daher wird vorgeschlagen, dass gute Umweltbedingungen paradoxerweise den Zelltod der Nekridien verursachen, um den Fadenbruch und damit die Reproduktion zu fördern.[32]
Stoffwechsel
Die Gattung Beggiatoa besteht hauptsächlich aus chemolithotrophen, sulfid-oxidierenden Bakterien. Die Palette der möglichen Stoffwechselwege ist jedoch sehr vielfältig und reicht von der Heterotrophie bis zur Chemolithoautotrophie. Aufgrund dieser großen Variabilität können sich die verschiedenen Arten und Stämme dieser Gattung stark voneinander unterscheiden.[27]
Kohlenstoff-Stoffwechsel
In der Gattung Beggiatoa gibt es sowohl autotrophe als auch heterotrophe Stoffwechselprozesse. Autotrophe Beggiatoa führen die CO2-Fixierung durch den Calvin-Zyklus und den Einsatz des Enzyms RuBisCO durch. Letzteres ist bei obligaten und fakultativen Autotrophen unterschiedlich stark reguliert. So kann RuBisCO im obligat autotrophen Stamm Beggiatoa sp. MS-81-1c[33] nicht unterdrückt werden, während es im fakultativ autotrophen Stamm Beggiatoa sp. MS-81-6[34] stark reguliert wird, um vom autotrophen zum heterotrophen Wachstum und umgekehrt zu wechseln. Neben den autotrophen Stämmen sind die meisten Süßwasser-Beggiatoa-Stämme heterotroph und benötigen organische Substrate für ihr Wachstum. Insbesondere können viele von ihnen als mixotroph angesehen werden, da sie heterotroph wachsen und organische Verbindungen oxidieren; aber auch Sulfid oder andere reduzierte Schwefelverbindungen als Elektronendonatoren nutzen können. Durch diese Strategie kann organischer Kohlenstoff für die Vermehrung der Biomasse gespeichert werde, eine autotrophe CO2-Fixierung ist dann nicht erforderlich. Mixotrophie wird als trophische Modalität für viele Süßwasserstämme zwar vermutet, wurde aber bisher nur in einem marinen Beggiatoa-Stamm, dem bezeichneten B. sp. MS-81-6, gefunden.[27][28] Auch ein Stoffwechselweg zur Nutzung von C1-Verbindungen (Moleküle mit einem einzigen Kohlenstoff-Atom) wurde im Stamm B. leptomitoformis D-402T, durch umfassende Analyse seiner Genomik, Physiologie und Biochemie/Molekularbiologie aufgedeckt.[17]
Stickstoff-Stoffwechsel
Die Beggiatoa-Bakterien zeigen eine große Vielseitigkeit bei der Verwertung von Stickstoffverbindungen. Stickstoff kann einerseits ein Wachstumsfaktor sein oder (wie beispielsweise als Nitrat) ein Elektronenakzeptor für die anaerobe Atmung. Heterotrophe Süßwasser-Beggiatoa-Arten assimilieren Stickstoff für ihr Wachstum. Zu den Stickstoffquellen gehören Nitrat, Nitrit, Ammoniak, Aminosäuren (wie Alanin , Asparagin und Asparaginsäure), Harnstoff und Thioharnstoff, je nach der Fähigkeit der einzelnen Arten und Stämme.[5][27]
Autotrophe vakuolierte Beggiatoa sind in der Lage, Nitrat in ihren Vakuolen in der 20.000-fachen Konzentration des umgebenden Meerwassers zu speichern und es unter anoxischen Bedingungen als terminalen Elektronenakzeptor zu nutzen.
Bei diesem Prozess, der als Dissimilatorische Nitratreduktion zu Ammonium (en.
, DNRA) bezeichnet wird, wird Nitrat zu Ammonium reduziert.
Die Fähigkeit, Nitrat als Elektronenakzeptor zu nutzen, ermöglicht die Besiedlung von anoxischen Umgebungen wie mikrobiellen Matten und Sedimenten. Mehrere Beggiatoa-Arten sind in der Lage, Stickstoff mit Hilfe des Enzyms Nitrogenase zu fixieren, z. B. Beggiatoa alba.[5][27]
Schwefel-Stoffwechsel
Eines der charakteristischen Merkmale der Gattung Beggiatoa ist die Produktion von intrazellulären Schwefeleinschlüssen, die aus der Oxidation reduzierter Schwefelquellen (z. B. Schwefelwasserstoff, H2S) resultieren. In autotrophen Beggiatoa-Vertretern ist Sulfid eine Energie- und Elektronenquelle für die Kohlenstofffixierung und das Wachstum. Die Oxidation von Sulfid kann aerob oder anaerob erfolgen, d. h. sie kann mit der Reduktion von Sauerstoff oder mit der Reduktion von Nitrat gekoppelt sein.
Der durch die Oxidation von Sulfid erzeugte Schwefel wird in internen Kügelchen („Globulen“, en.
, vgl. Granulen) gespeichert und kann wieder verwendet werden, wenn die Sulfidkonzentration sinkt.
Die vorübergehende Speicherung von elementarem Schwefel (S0) erhöht also die Anpassungsfähigkeit eines Organismus und seine Toleranz gegenüber Veränderungen der Sulfid- und Sauerstoffkonzentration.[5][27]
Aerobe Oxidation von Sulfiden:

Anaerobe Oxidation von Sulfiden:

Es gibt auch einige Fälle von Chemoorganotrophie. So betreibt beispielsweise der Stamm Beggiatoa sp. 35Flor[35] normalerweise eine aerobe mit der Oxidation von Sulfid gekoppelte Atmung; aber unter anoxischen Bedingungen wird eine andere Art der Atmung aktiviert: Die Energie wird dann chemoorganotroph aus der Oxidation von Polyhydroxyalkanoaten (PHA) gewonnen – das sind organische Verbindungen, die zuvor durch CO2-Fixierung während des chemolithotrophen Wachstums mit Hilfe von Sauerstoff und Sulfid synthetisiert wurden. In diesem Fall ist der Elektronenakzeptor der in der Zelle gespeicherte elementare Schwefel (S0), so dass das Endprodukt Schwefelwasserstoff ist.[23]
Anaerobe Atmung:

Wasserstoff-Stoffwechsel
Der Stamm B. sp. 35Flor ist auch in der Lage, Wasserstoff als alternativen Elektronendonator zu Sulfid zu nutzen. Dieser Oxidationsprozess kann Energie für Wartungs- und Assimilationszwecke liefern und kann den gespeicherten Schwefel zu reduzieren, wenn er übermäßig wird. Er kann allerdings den Organismen kein Wachstum verschaffen.[24]
Wasserstoff-Oxidation:

Phosphor-Stoffwechsel
Der Stoffwechsel von Beggiatoa umfasst die Verwendung von Phosphor in Form von Polyphosphat. Die Regulierung dieses Stoffwechsels hängt von den Umweltbedingungen ab. Eine sauerstoffreiche Umgebung führt zu einer Anhäufung von Polyphosphat, während Anoxie (in Verbindung mit einer steigenden Sulfidkonzentration) einen Abbau von Polyphosphat und seine anschließende Freisetzung aus den Zellen bewirkt. Das freigesetzte Phosphat kann sich dann als Phosphorit-Mineral in den Sedimenten ablagern, aber auch im Wasser gelöst bleiben.[27]
Ökologie und Anwendung
Wie beobachtet bilden Beggiatoa-Fadenbakterien dichte Matten auf Sedimenten in einer Vielzahl von Umgebungen, wo sie als weißliche Schicht erscheinen. Sie leben an der oxischen/anoxischen Grenzfläche, wo sie sowohl von der Anwesenheit von Schwefelwasserstoff (H2S) als auch von Sauerstoff (O2) profitieren. Die chemolithoautotrophen Stämme von Beggiatoa werden auch als wichtige Primärproduzenten in dunklen Umgebungen angesehen.[5]
Habitat
Aufnahme des DSRV Alvin, Bild der NOAA.
Die äußerst große Anpassungsfähigkeit der Stoffwechselvorgänge einzelner Beggiatoa-Vertreter in dieser Bakteriengattung ist eine evolutionäre Folge der außerordentlichen Variabilität der Umweltbedingungen, in denen sie leben. Beggiatoa ist fast benthisch, ihre Vertreter können einerseits in marinen Umgebungen (B. sp. MS-81-6 und B. sp MS-81-1c), andererseits auch in Süßwasserumgebungen (B. alba) vorkommen und benötigen nur Sulfid oder Thiosulfid (das Anion S=S– oder das Radikal S=S-, wiktionary: thiosulfide) als Elektronendonator und Oxidationsmittel. Sie sind in der Regel in Lebensräumen mit hohem Schwefelwasserstoffgehalt zu finden, z. B. in kalten Quellen, Schwefelquellen, mit Abwasser verunreinigtem Wasser, in Schlammschichten von Seen und in der Nähe hydrothermalen Schloten der Tiefsee. Beggiatoa findet man auch in der Rhizosphäre (Wurzelbereich) von Sumpfpflanzen,[36][13] im Boden, in Meeressedimenten und auch in Mangrovenlagunen, wo sie zum Lipid-Anteil der Sedimente beitragen.[37][38] Die Süßwasserarten haben typische Lebensräume in Schwefelquellen, Gräben, Pfützen, Feuchtgebieten, Seesedimenten und in Reisfeldern, wo sie in Verbindung mit den Wurzeln der Reispflanzen wachsen können. Die im Meerwasser lebenden Beggiatoa sind in Regionen zu finden, in denen ihre Energiequelle (Sulfid oder Thiosulfid) verfügbar ist. Sie können sowohl aus anorganischen als auch aus organischen Quellen gewonnen werden und sind in der Regel bei mikrooxischen Bedingungen, d. h. einer sehr niedrigen Sauerstoffkonzentration anzutreffen.[5] Außerdem findet man diese Gattung von Gammaprotobakterien auch in lokalisierten Bereichen anaerober Zersetzung, wie z. B. auf Kadavern im Wald und auf dem Meeresboden der Tiefsee.[5]

Vakuolisierte Beggiatoa (Beggiatoa mit sehr großen Vakuolen im Vergleich zum gesamten Zellvolumen) sind in küstennahen Auftriebsgebieten (z. B. an den Küsten Perus und Chiles), in hydrothermalen Schloten der Tiefsee und in Kaltwasserschloten (en.
) sehr häufig anzutreffen.
In diesen Umgebungen können Flockungsmatten wachsen und große Flächen bedecken, ihre haarähnlichen Filamente können eine Höhe von 30 cm über dem Substrat erreichen.
In hydrothermalen Schloten der Tiefsee und in
können Beggiatoa in Fäden wachsen, die einen Durchmesser von bis zu 200 mm (Mikrometern) haben, was sie zu den größten derzeit bekannten Prokaryoten macht.
Vakuolisierte Beggiatoa kommen auch in hypoxischen Meeresböden (mit wenig Sauerstoff) vor, wo die Filamente in den Sedimenten in einer Tiefe von etwa 2 bis 4 cm Zentimetern leben anzutreffen sind. In diesen Fällen können die bakteriellen Filamente von Beggiatoa der häufigste Teil der mikrobiellen Biomasse in den Sedimenten darstellen.[5]
Beggiatoa können auch in Salzsümpfen, Salinen und geothermisch aktiven Unterwasserhöhlen vorkommen. Einige Studien zu diesen Lebensräumen wurden in den Unterwasserhöhlen des dolomitischen Kalksteins in Capo Palinuro, Provinz Salerno (Italien) durchgeführt. Hier gibt es hydrothermale sulfidische Quellen, und der mikrobielle Biofilm steht in Verbindung mit den Bewegung dieser hydrothermalen Flüssigkeiten infolge der Gezeiten. Die in den Höhlen gefundenen Filament-Matten bestanden größtenteils aus Bakterien, die Beggiatoa, der verwandten Gattung Thiothrix, sowie Flexibacter (Klasse Cytophagia der Bacteroidetes) ähneln. Diese Beggiatoa-ähnlichen Filamente sind morphologisch denen ähnlich, die an Felsen und am Byssus der Muscheln aus den Hydrothermalquellen von Lucky Strike auf dem Mittelatlantischen Rücken gefunden wurden.[5]
Wechselwirkung mit anderen Organismen

Häufig können die Mikroorganismen der Gattung Beggiatoa komplexe mikrobielle Matten bilden, in denen sie zusammen mit vielen anderen fadenförmigen Bakterien, z. B. Cyanobakterien, leben. Die Cyanobakterien besetzen in der Regel die Oberflächenschicht der Matte und produzieren tagsüber eine große Menge an Sauerstoff, der aus der Photosynthese stammt.[5] Umgekehrt wachsen die Beggiatoa-Bakterien entlang der oxischen/anoxischen Grenzfläche zwischen Sauerstoff und Sulfid, d. h. unterhalb der phototrophen Mikroorganismen, wo sie (aufgrund des von ihnen akkumulierten elementaren Schwefels) weiße Flecken erzeugen.[5]
Nebenstehend eine Konzeptionszeichnung der Biomasseverteilung, nachdem sich die Organismen an unterschiedlich vorgegebene Lichtbedingungen angepasst haben („photische Zonen“), gemessen bei einer Bestrahlungsstärke der einfallenden Photonen von 1000 μmol/m²s (Mikromol Photonenpro Quadratmeter und Sekunde).[39] Die Oberflächenschichten der mikrobiellen Matte werden von dichten Populationen beweglicher Cyanobakterien (Microcoleus sp. und anderer fadenförmiger Arten) und fadenförmigen sulfidoxidierenden Bakterien (Beggiatoa spp.) dominiert. Unterhalb der schmalen photischen Zone befindet sich eine – aufgrund von enthaltenem Eisen(II)-sulfid (FeS) dunkle – Sedimentschicht. Im Detail:[39]
Rechts: Während der Dunkelakklimatisierung wird die gesamte Matte jedoch anoxisch, so dass die Beggiatoa an die Oberfläche der Matte wandern und eine weiße Schicht über den Cyanobakterien ausbilden. Damit entgehen sie den hohen Konzentrationen an Schwefelwasserstoff (H2S) und bleiben an der Sauerstoff/Sulfid-Grenzfläche (O2-H2S-Grenze), während die Cyanobakterien in einer dichten Schicht darunter blieben.[40][39]
Links: Bei schwachem Licht werden die oberen Schichten oxidiert und treiben die O2-H2S-Grenzfläche und damit Beggiatoa tiefer in die Matte, während sich die beweglichen Cyanobakterien bei den schwachen (nicht hemmenden) Lichtverhältnissen in Richtung der Mattenoberfläche bewegen.[39]
Mitte:Bei starkem Licht wird ein größerer Teil des Biofilms oxidiert, die Beggiatoa bewegen sich weiter nach unten; die beweglichen Cyanobakterien in den oberen Schichten beginnen sich ebenfalls etwas nach unten zu bewegen, um die hohen (für sie inhibitorischen) Lichtwerte an der Oberfläche zu vermeiden.[39]
Manchmal werden Beggiatoa-Matten auch durch Kieselalgen und grüne Euglenoiden angereichert.[41] Im Ökosystem des Guaymas-Beckens wurden auch noch andere Protisten wie Wimpertierchen und Dinoflagellaten in Verbindung mit den Matten gefunden; diese verzehren wahrscheinlich eine große Menge der bakterieller Biomasse.[42]
Da die mikrobiellen Matten bis zu 3 cm breit werden können, stellen sie eine Nahrungsquelle für viele Weidetiere dar. Ein Beispiel für einen Fressfeind ist das Wimpertierchen Colpidium, dessen Auftreten sogar ein Indikator für Wasserverschmutzung darstellt (siehe Colpidium §Indikator für die Wasserverschmutzung). Diese trophische Assoziation wurde auch in Mangrovensystemen beobachtet, wo Beggiatoa einen Teil der Meeressedimente bedecken. Diese Bakterien bilden dort einen wichtigen Beitrag zur Ernährung der Meiofauna, insbesondere von Rädertierchen (Rotifera), Vielborstern (Polychaeta), Fadenwürmern (Nematoda) und einigen Gruppen von Plattwürmern (Plathelminthes), Schlauchwürmern (hier: Aschelminthes) und Kiefermündchen (Gnathostomulida).[43] Es wurde eine bemerkenswerte Beziehung zwischen Nematoden und Beggiatoa festgestellt: Tatsächlich scheinen Nematoden die Entwicklung von Beggiatoa-Matten zu begünstigen, indem sie das Eindringen von Sauerstoff und die Diffusion von Nährstoffen in die Matte fördern.[44] Ein weiteres Beispiel für einen Fressfeind von Beggiatoa ist der blinde Höhlenkrebs Tethysbaena ophelicola (Ranzenkrebse) in der israelischen Ajalon-Höhle.
Außerdem sind viele Aasreste (Kadaver) mit Matten aus Beggiatoa-ähnlichen Fadenbakterien bedeckt, die anaerobe, sulfatreduzierende Bakterien überlagern. Die Kadaver locken an sich viele Aasfresser (Nekrophagen) an; aber wenn sie die Matte aufbrechen, wird Schwefelwasserstoff freigesetzt und vertreibt die Aasfresser.
Daher können Beggiatoa auch als ein „Schutz“ der Kadaver (en.
) vor den Aasfressern betrachtet werden.[45]
Rolle in biogeochemischen Kreisläufen

„Grotta sulfurea“, Capo Palinuro, Provinz Salerno
Mehrere Arten weißer Schwefelbakterien der Familie Beggiatoaceae (bzw. Thiotrichaceae) sind in der Lage, Nitrat (Nitrationen, NO3–), das aus flachen Küstensedimenten stammt und für den Stoffwechsel von grundlegender Bedeutung ist, zu akkumulieren, zu transportieren, sowie in ihren Fäden zu speichern. Die Reduktion von Nitrationen zu Ammonium setzt die Oxidation von Schwefelwasserstoff (H2S) voraus. Dieses Sulfid stammt außer bei geothermischen Quellen aus dem darunter liegenden anaeroben Sediment – in diesem Sediment findet eine dissimilatorische Sulfatreduktion statt,[41] die zur Bildung einer suboxischen Zone führt, die durch ein positives Redoxpotential und nur geringe Konzentrationen von freiem H2S gekennzeichnet ist. In der Meeresumwelt sid diese Arten wichtig, da sie eine grundlegende Rolle bei der Regulierung der H2S- und NO3–- Menge spielen.
- Einerseits ist die Regulierung der Konzentration von freiem H2S in Meeressedimenten von grundlegender Bedeutung, da sulfidarme Oberflächensedimente für das Überleben der benthischen Fauna von entscheidender Bedeutung sind (Sulfid bzw. Schwefelwasserstoff ist für die Bodenfauna und andere im Sediment lebende Organismen hochgiftig)
- Andererseits ist die NO3–-Reduktion wichtig für die Kontrolle der Eutrophierung in stickstoffbegrenzten (stickstoffarmen) Küstengewässern.[46]
Die Tatsache, dass Beggiatoa in der Lage ist, Phosphor als Polyphosphat zu akkumulieren und anschließend unter anoxischen Bedingungen als Phosphat in die Wassersäule freizusetzen, kann die Verfügbarkeit von Phosphor für Produzent (Ökologie)Primärproduzenten erhöhen. Einige Studien über den Phosphorkreislauf und die Freisetzung von Phosphor in Verbindung mit Beggiatoa wurden in der Ostsee durchgeführt. Diese Studien haben gezeigt, dass die Reduktion von Sulfid durch diese Bakterien die Geschwindigkeit der Eisensulfidbildung in den Sedimenten verringern und somit die Fähigkeit des Sediments, Phosphor zu binden, erhöhen kann.[27]
Anwendungen
Da Beggiatoa in verschmutzten Meeresumgebungen vorkommen und gedeihen, können sie als Bioindikator angesehen werden;[47] genauso wie ihre Fressfeinde wie das Wimpertierchen Colpidium.
Beggiatoa und andere verwandte Fadenbakterien können Absetzprobleme in kommunalen Kläranlagen wie in industriellen Abwasserkläranlagen (etwa bei der Konserven-Herstellung, Papierherstellung, Brauerei und Müllerei) verursachen, was zu dem Phänomen des „
“ ([en]) führt.
Geeignete Beggiatoa-Stämme sind auch in der Lage, Schwefelwasserstoff im Boden zu entgiften und spielen eine Rolle bei der Immobilisierung von Schwermetallen.[41][48]
Kultivierung
Selektive Anreicherung
Die erfolgreichsten Anreicherungen für Beggiatoa spp. wurden mit Hilfe einer flachen Schale oder eines Aquariums vorgenommen, in das einige Zentimeter Sand, verschiedene Mengen an Kalziumsulfat (CaSO4) und Dikaliumhydrogenphosphat (K2HPO4), eine Quelle komplexer organischer Polymere (Biopolymere) wie Algen, einige Zentimeter sulfidreicher Meeresschlamm und Meerwasser gegeben wurden. Die Anreicherung muss die richtige Sulfid-Sauerstoff-Grenzfläche aufweisen, was nur möglich ist, wenn Luft zugeführt wird, zum Beispiel durch einen langsamen, stetigen Fluss von frisch belüftetem Meerwasser.[5]
Eine andere Art der Anreicherung von Beggiatoa spp. basiert auf der Verwendung von extrahiertem getrocknetem Gras oder Heu in einem mineralischen Medium, da die komplexen Polymere wie Zellulosereste in diesem Material für die Sulfatreduktion sorgen. Dies liefert auch den Schwefelwasserstoff, der für die Anreicherung von Beggiatoa notwendig ist.[5]
Isolierung von Reinkulturen
Es gibt drei verschiedene Techniken, um isolierte Beggiatoa-Stämme in Reinkultur zu erhalten:
- Isolierung auf Agarplatten (Petrischalen)
- Isolierung in Flüssigmedien
- Isolierung und Kultivierung in Gradientenmedien (Medium mit Konzentrationsgefälle)
Isolation auf Agarplatten
Dieses Verfahren zur Gewinnung eines isolierten heterotrophen Stammes erfordert eine Agarplatte mit verdünntem organischem Substrat, z. B. einer kleinen Menge Pepton. Dann müssen Büschel von Beggiatoa-Fäden aus der Umgebung gesammelt, mit steriler Waschlösung gewaschen und auf die Agarplatte gelegt werden (Inokulation). Auf diese Weise entstehen einige wachsende Fäden, die sich von dem zentralen Inokulum entfernen und als Inokulum für eine neue Agarplatte verwendet werden können. Für die Isolierung mariner Beggiatoa-Stämme (die autotrophes Wachstum zeigen) ist es für diese obligat mikroaereophile Organismen unerlässlich, mikrooxische Bedingungen (mit ein wenig Sauerstoff) zu schaffen und spezielle Agarplatten zu verwenden, die aus gefiltertem Meerwasser mit Natriumsulfid (Na2S) und Natriumacetat (NaC(O)CH3) hergestellt werden. Im Gegensatz dazu muss die Isolierung der Süßwasserstämme unter oxischen Bedingungen (Luftatmosphäre) erfolgen, wobei verschiedene Medien mit einer niedrigen Konzentration einer einzelnen organischen Verbindung wie Na2S, Acetat, oder Thiosulfat (S2O32−) verwendet werden.[5]
Isolation in Flüssigmedien
Flüssigmedien werden häufig zur Anreicherung, Titrierung (MPN-Auszählung,
) und Massenkultivierung von Beggiatoa verwendet.
Für eine erfolgreiche Kultivierung von heterotrophen oder mixotrophen Süßwasser-Beggiatoa müssen Flüssigmedien geringe Mengen an Kohlenstoffsubstrat enthalten, entweder Bodenextrakte oder Acetat. Der Referenzstamm der Typusart (B. alba str. B18LD) und verwandte Stämme werden im Allgemeinen in Medien kultiviert, die eine Salzbasis, Acetat als Kohlenstoffquelle und variable Hefeextrakt- und Sulfidzusätze enthalten.[49] Einige marine autotrophe Beggiatoa-Stämme werden auch auf definierten flüssigen mineralischen Medien mit Thiosulfat, CO2 unter mikro-oxischen Bedingungen – Belüftung mit 0,25 % O2 (v/v) in der Gasphase – kultiviert.[5]
Isolation und Kultivatierung in Gradienten-Medien
Autotrophe Stämme, die aus einer Einzelfilamentisolierung auf Agar stammen, können leicht in Sulfidgradientenröhrchen (Röhrchen mit Konzentrationsgefälle ans Sulfid, vgl. auch Winogradsky-Säule) gehalten und vermehrt werden. Dabei werden sulfidreiche Agarpfropfen mit sulfidfreiem Weichagar überlagert. Die Röhrchen sind locker verschlossen, um den Austausch von Gasen oben im Kopfraum mit der Atmosphäre zu ermöglichen. Dadurch bilden sich zwei vertikal gegenüberliegende Schichten, von denen die eine Sulfid und die andere Sauerstoff enthält: Dies ermöglicht das Wachstum einer gut definierten Beggiatoa-Schicht an der sich ausbildenden Sulfid-Sauerstoff-Grenzfläche. Der Aufbau des Gradientenmediums erfordert unterschiedliche Mengen an J3-Medium – hergestellt aus Agar und Natron (NaHCO3) – das mit neutralisiertem Natriumsulfid (Na2S) ergänzt wird und in einem Schraubverschlussröhrchen untergebracht ist. Hier wird die Schwefelquelle durch den Sulfidfluss bereitgestellt. Eine weitere „Schicht“ besteht aus NaHCO3 ohne Sulfid oder Thiosulfat: Das gesamte Sulfid befindet sich unterhalb der Grenzfläche zwischen dem sulfidischen Agarpfropfen und dem übergelagerten sulfidfreien Agar, während sich im oberen Teil des Röhrchens eine weitere Schicht befindet, die das Sauerstoffreservoir darstellt. Aufgrund der Reaktion zwischen Sulfid und Sauerstoff beginnt sich ein Gefälle zu bilden: die Fäden vermehren sich schnell an der Sulfid-Sauerstoff-Grenzfläche und bilden eine markante Schicht oder „Platte“ mit einer Stärke von ca. 1 mm. Man sehen, dass diese Bakterien der Grenzfläche folgen und aufgrund der allmählichen Erschöpfung des Sulfid-Reservoirs langsam absteigen.[5]
Siehe auch
- Ajalon-Höhle §Speläobiologie
- Cueva de Villa Luz §Höhlenökologie
- Muchimuk-Höhlensystem §Die Opal-Speläotheme des Höhlensystems
Literatur und Weblinks
- MicrobeWiki: Beggiatoa, Kenyon College (kenyon.edu), Department of Biology
- OneZoom: Beggiatoa
- Beggiatoa and hydrocarbon seeps, Kapitel: Roxanne L. Nikolaus: Unique bacteria thriving in a unique environment". Memento im Webarchiv vom 11. August 2014.
- Lars G. Ljungdahl, Michael W. Adams, Larry L. Barton, James G. Ferry, Michael K. Johnson: Biochemistry and Physiology of Anaerobic Bacteria, Springer-Verlag New York, 2003. ISBN 978-0-387-95592-6. siehe insbes. S. 17.
Einzelnachweise
- ↑ Vittore Benedetto Antonio Trevisan: Prospetto della Flora Euganea (Prosp. Fl. Eugan.). In: Coi Tipi Del Seminario 1842, S. 1–68. Full Text, hier: S. 55f – „Beggiatoa“. Epub 29. April 2008. (italienisch)
- ↑ a b Lars G. Ljungdahl: Biochemistry and physiology of anaerobic bacteria. Springer, 2003, ISBN 978-0-387-95592-6, S. 17.
- ↑ a b Pratap N. Mukhopadhyaya, Chirajyoti Deb, Chandrajit Lahiri, Pradosh Roy: A soxA gene, encoding a diheme cytochrome c, and a sox locus, essential for sulfur oxidation in a new sulfur lithotrophic bacterium. In: ASM Journals: Journal of Bacteriology, Band 182, Nr. 15, August 2020, S. 4278–4287; doi:10.1128/JB.182.15.4278-4287.2000, PMID 10894738, PMC 101942 (freier Volltext), Epub 20. Dezember 2020.
- ↑ a b William R. Strohl, John M. Larkin: Cell division and trichome breakage in Beggiatoa. In: Current Microbiology, Band 1, Nr. 3, 1978, S. 151—155; doi:10.1007/BF02601668 , PMID 23338140, Epub. 8. April 2007.
- ↑ a b c d e f g h i j k l m n o p q r s t Andreas Teske, Douglas C. Nelson: The Genera Beggiatoa and Thioploca. In: M. Dworkin, S. Falkow, E. Rosenberg, K. H. Schleifer (Hrsg.): The Prokaryotes, Band 6: Proteobacteria: Gamma Subclass, Springer, New York, NY, 2006, S. 784–810, Print ISBN 978-0-387-25496-8; doi:10.1007/0-387-30746-x_27, PDF, Online ISBN 978-0-387-30746-6, Epub 1. April 2021.
- ↑ a b c d e f g Aidan C. Parte, J. P. Euzéby (LPSN): Search: Beggiatoa und Beggiatoa Trevisan 1842
- ↑ Galina Dubinina, Alexander Savvichev, Maria Orlova, Ekaterina Gavrish, Suzanne Verbarg, Margarita Grabovich: Beggiatoa leptomitoformis sp. nov., the first freshwater member of the genus capable of chemolithoautotrophic growth. In: International Journal of Systematic and Evolutionary Microbiology, Microbiology Society, Band 67, Nr. 2, 1. Februar 2017, S. 197–204; doi:10.1099/ijsem.0.001584, PMID 27902215.
- ↑ Marc Mußmann, Fen Z. Hu, Michael Richter, Dirk de Beer, André Preisler, Bo B. Jørgensen, Marcel Huntemann, Frank Oliver Glöckner, Rudolf Amann, Werner J. H. Koopman, Roger S. Lasken, Benjamin Janto, Justin Hogg, Paul Stoodley, Robert Boissy, Garth D. Ehrlich: Insights into the genome of large sulfur bacteria revealed by analysis of single filaments. In: PLOS Biology. 5, Nr. 9, September 2007, S. e230. doi:10.1371/journal.pbio.0050230. PMID 17760503. PMC 1951784 (freier Volltext).
- ↑ a b A. Fomenkov. T. Vincze, M. Y. Grabovich, G. Dubinina, M. Orlova, E. Belousova, R. J. Roberts: Complete Genome Sequence of the Freshwater Colorless Sulfur Bacterium Beggiatoa leptomitiformis Neotype Strain D-402T. In: Genome announcements, Band 3, Nr, 6, 2015; doi:10.1128/genomeA.01436-15, PMID 26659680.
- ↑ Hans Reichenbach: Taxonomy of the gliding bacteria. In: Annual Review of Microbiology. 35, Nr. 1, 1. Oktober 1981, S. 339–364. doi:10.1146/annurev.mi.35.100181.002011. PMID 6794424.
- ↑ David A. Stahl, David J. Lane, Gary J. Olsen, Debra J. Heller, Thomas M. Schmidt, Norman R. Pace: Phylogenetic Analysis of Certain Sulfide-Oxidizing and Related Morphologically Conspicuous Bacteria by 5S Ribosomal Ribonucleic Acid Sequences. In: International Journal of Systematic and Evolutionary Microbiology. 37, Nr. 2, 1. April 1987, ISSN 1466-5026, S. 116–122. doi:10.1099/00207713-37-2-116.
- ↑ a b c d e NCBI: *beggiatoa (Namensabkömmlinge), Beggiatoa, Beggiatoa Trevisan 1842 (genus); graphisch: Beggiatoa, auf: Lifemap NCBI Version.
- ↑ a b c Azeem Ahmad, James P. Barry, Douglas C. Nelson: Phylogenetic affinity of a wide, vacuolate, nitrate-accumulating Beggiatoa sp. from Monterey Canyon, California, with Thioploca spp. In: ASM Journals: Applied and Environmental Microbiology, Band 65, Nr. 1, Januar 1999, S. 270–277, bibcode:1999ApEnM..65..270A; doi:10.1128/AEM.65.1.270-277.1999, PMID 9872789, PMC 91012 (freier Volltext), Epub 17. Dezember 2020.
- ↑ David R. Boone, Don J. Brenner, Richard W. Castenholz, Paul De Vos, George M. Garrity, Noel R. Krieg, M. Goodfellow: Bergey's manual of systematic bacteriology, 2. Auflage, Springer, New York 2001–2012, ISBN 978-0-387-21609-6, OCLC 619443681.
- ↑ a b c WoRMS: Beggiatoa V.B.A.Trevisan, 1845
- ↑ a b c AlgaeBase: Beggiatoa Trevisan, 1845
- ↑ a b Maria V. Orlova, Sergey V. Tarlachkov, Eugenia I. Kulinchenko, Galina A. Dubinina, Maria N. Tutukina, Margarita Y. Grabovich: Genomics and Biochemistry of Metabolic Pathways for the C1 Compounds Utilization in Colorless Sulfur Bacterium Beggiatoa leptomitoformis D-402. In: Indian Journal of Microbiology, Band 58, Nr. 4, Dezember 2018, S. 415–422; doi:10.1007/s12088-018-0737-x, PMID 30262951, PMC 6141403 (freier Volltext), Epub 2. Mai 2018.
- ↑ NCBI: unclassified Beggiatoa (list)
- ↑ NCBI: txid2587807[Organism:noexp]
- ↑ NCBI: txid2587806[Organism:noexp]
- ↑ a b NCBI: Beggiatoa sp. 'Orange Guaymas'
- ↑ a b c Matthias Winkel, Verena Salman-Carvalho, Tanja Woyke, Michael Richter, Heide N. Schulz-Vogt, Beverly E. Flood, Jake V. Bailey, Marc Mußmann: Single-cell Sequencing of Thiomargarita Reveals Genomic Flexibility for Adaptation to Dynamic Redox Conditions. In: Front. Microbiol., Band 7, 21. June 2016; doi:10.3389/fmicb.2016.00964, ISSN 1664-302X. Siehe insbes. Fig. 1.
- ↑ a b Anne Schwedt, Anne-Christin Kreutzmann, Lubos Polerecky, Heide N. Schulz-Vogt: Sulfur respiration in a marine chemolithoautotrophic Beggiatoa strain. In: Frontiers in Microbiology: Microbial Physiology and Metabolism, Band 2, 9. Januar 2012, S. 276; doi:10.3389/fmicb.2011.00276, PMID 22291687, PMC 3253548 (freier Volltext).
- ↑ a b Anne-Christin Kreutzmann, Heide N. Schulz-Vogt: Oxidation of Molecular Hydrogen by a Chemolithoautotrophic Beggiatoa Strain. In: ASM Journals: Applied and Environmental Microbiology, Band 82, Nr. 8, S. 2527–2536, bibcode:2016ApEnM..82.2527K; doi:10.1128/AEM.03818-15, PMID 26896131, PMC 4959497 (freier Volltext)
- ↑ a b Julia Peixoto de Albuquerque, Carolina Neumann Keim, Ulysses Lins: Comparative analysis of Beggiatoa from hypersaline and marine environments. In: Micron, Band 41, Nr. 5, Juli 2010, S. 507–517; doi:10.1016/j.micron.2010.01.009 , PMID 20207153, Epub 11. Februar 2010.
- ↑ a b S. Grünke, A. Lichtschlag, D. de Beer, J. Felden, V. Salman, A. Ramette, H. N. Schulz-Vogt, A. Boetius: Mats of psychrophilic thiotrophic bacteria associated with cold seeps of the Barents Sea. In: Biogeosciences, Band 9, S. 2947–2960, 6. August 2012; doi:10.5194/bg-9-2947-2012, PDF.
- ↑ a b c d e f g h i j k Matti Ruuskanen: The genus Beggiatoa and its effects on the nutrient cycles of the Baltic Sea. Thesis (Bachelor of Science), University of Turku, Mai 2014, Epub 8. April 2016.
- ↑ a b c K. D. Hagen, D. C. Nelson: Use of Reduced Sulfur Compounds by Beggiatoa spp.: Enzymology and Physiology of Marine and Freshwater Strains in Homogeneous and Gradient Cultures. In: ASM Journals Applied and Environmental Microbiology, Band 63, Nr. 10, Oktober 1997, S. 39573964; doi:10.1128/aem.63.10.3957-3964.1997 , PMID 16535709, PMC 1389265 (freier Volltext), Epub 31. Januar 2021.
- ↑ Lotte Burkhardt 2022: Eine Enzyklopädie zu eponymischen Pflanzennamen: Von Menschen & ihren Pflanzen – Berlin: Botanic Garden and Botanical Museum Berlin, Freie Universität Berlin 2022. – doi:10.3372/epolist2022.
- ↑ Terme Euganee, auf TripAdvisor
- ↑ Euganeische Thermen — Kultur und Wellness, auf Visit Abano Montegrotto
- ↑ a b Anja Kamp, Hans Røy, Heide N. Schulz-Vogt: Video-supported analysis of Beggiatoa filament growth, breakage, and movement. In: Microbial Ecology, Band 56, Nr. 3, Oktober 2008, S. 484–491; doi:10.1007/s00248-008-9367-x, PMID 18335158, PMC 2755761 (freier Volltext), Epub 12. März 2008.
- ↑ NCBI: Beggiatoa sp. MS-81-1c (species)
- ↑ NCBI: Beggiatoa sp. MS-81-6 (species)
- ↑ NCBI: Beggiatoa sp. 35Flor (species)
- ↑ Michael Dudley: Beggiatoa. Soil Microbiology –BIOL/CSES 4684. Virginia Tech (vt.edu). Archiviert vom Original am 7. Februar 2009.
- ↑ A. SamKamaleson, Maria-Judith Gonsalves: Role of sulfur-oxidizing bacteria on the ecology in tropical mangrove sediments. In: Regional Studies in Marine Science. 28, April 2019, ISSN 2352-4855, S. 100574. doi:10.1016/j.rsma.2019.100574.
- ↑ Maïtena R. N. Jean, Silvina Gonzalez-Rizzo, Pauline Gauffre-Autelin, Sabine K. Lengger, Stefan Schouten, Olivier Gros: Two New Beggiatoa Species Inhabiting Marine Mangrove Sediments in the Caribbean. In: PLOS ONE, Band 10, Nr. 2, 17. Februar 2015, S. e0117832, bibcode:2015PLoSO..1017832J; doi:10.1371/journal.pone.0117832, PMID 25689402, PMC 4331518 (freier Volltext).
- ↑ a b c d e André Preisler, Dirk de Beer, Anna Lichtschlag, Gaute Lavik, Antje Boetius, Bo Barker Jørgensen: Biological and chemical sulfide oxidation in a Beggiatoa inhabited marine sediment. In: Nature ISME Journal, Band 1, August 2007, S. 341–353; doi:10.1038/ismej.2007.50, Epub 28. Juni 2007.
- ↑ Mads Lichtenberg, Paulo Cartaxana, Michael Kühl: Vertical Migration Optimizes Photosynthetic Efficiency of Motile Cyanobacteria in a Coastal Microbial Mat. In: Frontiers in Marine Science. 7, 25. Mai 2020, ISSN 2296-7745. doi:10.3389/fmars.2020.00359. Reihe: Advances and Challenges in Microphytobenthos Research: From Cell Biology to Coastal Ecosystem Function.
- ↑ a b c Tom Fenchel, Catherine Bernard: Mats of colourless sulphur bacteria. I. Major microbial processes. In: Marine Ecology Progress Series. 128, 23. November 1995, ISSN 0171-8630, S. 161–170. bibcode:1995MEPS..128..161F. doi:10.3354/meps128161. PDF.
- ↑ Alexis Pasulka, Sarah K. Hu, Peter D. Countway, Kathryn J. Coyne, Stephen C. Cary, Karla B. Heidelberg, David A. Caron: SSU-rRNA Gene Sequencing Survey of Benthic Microbial Eukaryotes from Guaymas Basin Hydrothermal Vent. In: The Journal of Eukaryotic Microbiology. 66, Nr. 4, Juli 2019, S. 637–653. doi:10.1111/jeu.12711. PMID 30620427.
- ↑ Pierre-Yves Pascal, Stanislas Dubois, Henricus T. S. Boschker, Olivier Gros: Trophic role of large benthic sulfur bacteria in mangrove sediment. In: Marine Ecology Progress Series. 516, 3. Dezember 2014, ISSN 0171-8630, S. 127–138. bibcode:2014MEPS..516..127P. doi:10.3354/meps11035. PDF.
- ↑ Humbert Salvadó, Alejandro Palomo, Meritxell Mas, Jaume Puigagut, Mª del Pilar Gracia: Dynamics of nematodes in a high organic loading rotating biological contactors. In: Water Research. 38, Nr. 10, Mai 2004, S. 2571–2578. doi:10.1016/j.watres.2004.03.007. PMID 15159160.
- ↑ Paul K. Dayton, John S. Oliver, Simon F. Thrush, Kamille Hammerstrom: Bacteria defend carrion from scavengers. In: Antarctic Science. 31, Nr. 1, Februar 2019, ISSN 0954-1020, S. 13–15. bibcode:2019AntSc..31...13D. doi:10.1017/S0954102018000457.
- ↑ Mikio Sayama, Nils Risgaard-Petersen, Lars Peter Nielsen, Henrik Fossing, Peter Bondo Christensen: Impact of bacterial NO3- transport on sediment biogeochemistry. In: Applied and Environmental Microbiology. 71, Nr. 11, November 2005, S. 7575–7577. doi:10.1128/AEM.71.11.7575-7577.2005. PMID 16269807. PMC 1287653 (freier Volltext).
- ↑ Joel K. Elliott, Erin Spear, Sandy Wyllie-Echeverria: Mats of Beggiatoa bacteria reveal that organic pollution from lumber mills inhibits growth of Zostera marina. In: Marine Ecology, Band 27, Nr. 4, 21. November 2006, S. 372–380, bibcode:2006MarEc..27..372E, ISSN 1439-0485; doi:10.1111/j.1439-0485.2006.00100.x.
- ↑ Lukasz Drewniak, Pawel S. Krawczyk, Sebastian Mielnicki, Dorota Adamska, Adam Sobczak, Leszek Lipinski, Weronika Burec-Drewniak, Aleksandra Sklodowska: Physiological and Metagenomic Analyses of Microbial Mats Involved in Self-Purification of Mine Waters Contaminated with Heavy Metals. In: Frontiers in Microbiology. 7, 10. August 2016, S. 1252. doi:10.3389/fmicb.2016.01252. PMID 27559332. PMC 4978725 (freier Volltext).
- ↑ Thomas M. Schmidt Boaz Arieli, Yehuda Cohen, Etana Padan, William R. Strohl: Sulfur metabolism in Beggiatoa alba. In: ASM Journals: Journal of Bacteriology, Bamd 169, Nr. 12, Dezember 1987, S. 5466–5472; doi:10.1128/jb.169.12.5466-5472.1987, PMID 3316186, PMC 213973 (freier Volltext), Epub 2. Februar 2021. PDF.

{kind=link}
{kind=link}
{kind=link}