Nucleocytoviricota
| Nucleocytoviricota | ||||||||
|---|---|---|---|---|---|---|---|---|

EM-Aufnahme eines Virions (Virusteilchens) des | ||||||||
| Systematik | ||||||||
| ||||||||
| Taxonomische Merkmale | ||||||||
| ||||||||
| Wissenschaftlicher Name | ||||||||
Nucleocytoviricota
| ||||||||
| Kurzbezeichnung | ||||||||
| NCLDV | ||||||||
| Links | ||||||||
|
Das Phylum der Nucleocytoviricota[1] (früher auch englisch Nucleocytoplasmic large DNA viruses, NCLDV) umfasst eine heterogene Gruppe meist großer dsDNA-Viren, die eine Reihe bestimmter Gene (NCLDV
) aufweisen, die gewöhnlichen Viren fehlen. Gründungsmitglieder der Gruppe (2001) sind die Familien
Asfarviridae, Iridoviridae und Poxviridae (Pockenviren), sowie die Phycodnaviridae. Weitere Mitglieder sind die Familien Mimiviridae (synonym mit „Megaviridae“), Marseilleviridae, Ascoviridae. Vorgeschlagene Mitglieder sind die Pithoviren, Orpheoviren und Pandoraviren, gegebenenfalls mit jeweils eigene Familien,[4] sowie etliche mehr. Mögliche Kandidaten sind Dinodnavirus, „Medusavirus“,[5][6][7][8][9] „Urceolovirus“[10] und „Yaravirus“.[11][12]
Diese Mitglieder bilden – wie inzwischen mehrfach bestätigt – eine monophyletische Verwandtschaftsgruppe, d. h. sie haben einen gemeinsamen viralen Vorfahren.[13][14][15] Man hatte daher zunächst verschiedentlich vorgeschlagen, diese Gruppe als „Megavirales“ in den Rang einer neuen Virusordnung zu erheben.[16] Nachdem das International Committee on Taxonomy of Viruses (ICTV) seit 2018 auch Ränge oberhalb dem der Ordnung zuließ,[17][18] wurden aufgrund der hohen Diversität dieser Gruppe solche höheren Ränge favorisiert. Damit wurde es ermöglicht, die Hauptgruppen der NCLDV als Ordnungen oder gar Klassen aufzustellen.[14] So schlugen beispielsweise Guglielmini et al. (2019) die erweiterte Familie Mimiviridae auf als eine Ordnung und schlugen dafür die nun frei gewordene Bezeichnung „Megavirales“ vor.[19]
Diese Entwicklung erreichte 2019/2020 mit der
Nummer 35 des ICTV ihren vorläufigen Abschluss mit der offiziellen Anerkennung der NCLDV als Phylum Nucleocytoviricota (ursprünglich „Nucleocytoplasmaviricota“[2]) mit den Klassen Megaviricetes und Pokkesviricetes durch das ICTV im März 2020. Die Ordnung der erweiterten Mimiviridae heißt jetzt Imitervirales.[1]
Genom
Das Genom der NCLDV ist vielfältig und hat eine Größe im Bereich 100 bis über 1500 kb („Tupanvirus“). Viele NCLDV sind damit Riesenviren (englisch giant viruses, giruses), wofür meist eine Überschreitung der 300 kb-Grenze vorausgesetzt wird (bei Yutin und Koonin von 500 kb[14][20]). Umgekehrt sind alle Riesenviren (bisher) dsDNA-Viren im Phylum NCLDV. Die größten Vertreter bilden größere Viruspartikel und haben ein größeres Genom als viele kleine Bakterien – die Mimiviren wurden zunächst sogar für (parasitäre) Bakterien gehalten, zumal sie auch Gram-Färbung zeigen. Zur späten Entdeckung der meisten Riesenviren (zahlreich erst ab etwa 2003) trug bei, dass sie bei der Suche nach Viren in den Filtern (mit typischer Porengröße von 0,2 μm) hängen blieben, die Bakterien und Protisten von Viren abtrennen sollten, langsamer zu leicht sichtbaren Klumpen aggregieren und sich auch langsamer vermehren.[21] 2020 berichteten Wissenschaftler Stoffwechselgene in den metabolismuslosen Viren gefunden zu haben, was darauf hindeutet, dass diese den Metabolismus ihrer Wirtszellen ändern.[22][23]
Wirte
Obwohl die NCLDV mehr eigene Proteine kodieren als gewöhnliche Viren (oft Hunderte statt kaum ein Dutzend), sind sie wirtspezifisch. Unter den Wirten sind ausschließlich komplex-zelluläre Organismen (Eukaryoten): viele Protisten (z. B. Amöben und Algen), aber auch Wirbeltiere und Insekten.[21]
Ursprünglich sechs, nun neun gemeinsame (homologe) Gene sind in allen NCLDV zu finden (NCLDV
), 177 weitere Gene (Stand 2009)[24] kommen in mindestens zwei der Familien vor. Dazu gehören vier Gene, welche die DNA-Replikation und Reparatur-Mechanismen betreffen:
die DNA-Polymerase-Familie B, die Topoisomerase II A, die „Flap“-Endonuklease und das Ringklemmenprotein (Proliferating-Cell-Nuclear-Antigen) sowie die RNA-Polymerase II und den Transkriptionsfaktor II B. Die Gene mancher NCLDV enthalten auch Introns.
Vermehrungszyklus
Manche der NCLDV vermehren sich ganz oder teilweise im Zellplasma (Zytoplasma) der eukaryotischen Wirtszellen, andere gehen möglicherweise durch eine nukleare Phase im Zellkern.[14] Viele NCLDV bilden nach der Infektion im Zytoplasma ihres Wirtes eine mikroskopisch sichtbare Produktionsstätte (Virusfabrik, englisch virus factory) aus. Bei diesen gibt es zum Teil andere Viren, die deren Syntheseapparat für ihre eigene Vermehrung nutzen und daher Virophagen (‚Virenfresser‘) genannt werden. Der erste entdeckte Fall dieser Art war Sputnikvirus 1 mit dem NCLDV Mamavirus, weitere Virophagen wie ‚Zamilon‘ befallen ebenfalls Vertreter der Mimiviridae. [25] Zwar werden Viren traditionell und meist auch heute noch nicht als Lebewesen angesehen, die durch NCLDV vollbrachten Leistungen verringern aber die Kluft zur belebten Welt. Unverändert gültig ist, dass Virionen keinen Stoffwechsel aufweisen, der auf ATP als „Energiewährung“ beruht.
Horizontaler Gentransfer
Zwischen den Riesenviren als viralen Endocytobionten (Organismen, die in den Zellen anderer Organismen leben oder sich vermehren)[26] und ihren oft amöboiden Wirten lässt sich massiver horizontaler Gentransfer in beiden Richtungen (AtoV, Amöbe auf Virus und umgekehrt VtoA, Virus auf Amöbe) nachweisen.[5]
Die Tatsache, dass für einige der unter den NCLDV verbreiteten Genen keine Entsprechung in zellulären Organismen gefunden wurde, wurde vom Team um Didier Raoult als Hinweis auf eine vierte Domäne des Lebens gedeutet, deren Vertreter bis auf die parasitierenden NCLDV ausgestorben sei. Diese
-Hypothese ist nicht mehr haltbar, seit für mehrere in NCLDV-Gruppen verbreitete Gene Abstammungen von weit auseinander liegenden Stellen im Baum des Lebens gezeigt wurden, überwiegend von verschiedenen Eukaryonten, aber auch von Bakterien.[27][28] Allerdings beflügelt die Entdeckung jeder neuer Familie der Riesenviren die Diskussion von Neuem, wie das Beispiel der Medusaviridae zeigt.[5][9]
Eine Zusammenfassung dieser Diskussion findet sich bei Traci Watson (2019).[29]
Es wurde in diesem Zusammenhang sogar nachgewiesen, dass es nicht nur einen Gentransfer zwischen den amöboiden Wirten und den Riesenviren als viralen Endocytobionten gibt (VtoA/AtoV), sondern sogar zwischen den Viren und gleichzeitig vorhandenen bakteriellen Endocytobionten.[30] Möglicherweise könnte dies die merkwürdige Übereinstimmung im Abwehrsystem CRISPR der Bakterien gegenüber Viren (also Bakteriophagen) einerseits und dem Abwehrsystem MIMIVIRE der Mimiviren gegenüber parasitierenden Satellitenviren (also Virophage)[25] andererseits erklären.
Endogenisierung
Die Landpflanzen Physcomitrella patens (Laubmoose) und der Selaginella moellendorffii (Lycophyten) besitzen offene Leserahmen (
), die von endogenisierten NCLDVs einer noch unbekannten Familie stammen könnten.[31]
Histone
Eine Reihe von NCLDVs kodiert Histone oder Histon-Homologe.
Beispiele finden sich in den Familien Marseilleviridae, Phycodnaviridae mit „Pandoravirus“ und „Clandestinovirus“, Iridoviridae, „Medusaviridae“ und Unterfamilie „Klosneuvirinae“. Anderweitige Beispiele gibt es in der Gattung Bracovirus (alias Bracoviriform, Spezies „
“, CvBV, zu NCLDV?)[32] und der Familie Nudiviridae.2022 wurde die Rolle von Histonen im Replikationszyklus einer Reihe von Viren umfassend untersucht.[33]
Systematik
Innere Systematik und Forschungsgeschichte
Trotz ihrer Diversität bilden die NCLDV wie mehrfach bestätigt eine Verwandtschaftsgruppe (Taxon),[34] wobei die Riesenviren von (verschiedenen) Gruppen kleinerer NCLDV abzustammen scheinen, statt umgekehrt.[35]
- „Irido-Ascoviridae“: Mehrere Studien unterstützen seit dem Jahr 2000 die Annahme, dass die Ascoviridae sich aus den Iridoviridae entwickelt haben,[36][37][38][39] es könnte aber auch umgekehrt sein.[40][41] Weiter wird vermutet, dass sich aus den Ascoviridae die Gattung Ichnovirus (Familie Polydnaviridae) entwickelt hat.[38]
- Dinodnavirus: Untersuchungen des Genoms von Heterocapsa circularisquama DNA virus 01 (Gattung Dinodnavirus) haben 2009 gezeigt, dass diese ursprünglich in die Familie Phycodnaviridae gestellte Gattung eher zur Familie der Asfarviridae gehört.[42]
- „Urceolovirus“: Eine weitere Spezies von DNA-Riesenviren, „Urceolovirus corneum“ („KLaHel“), erstmals berichtet 2015, könnte Mitglied der NCLDV sein.[10][43][44][45]
- Klosneuviren: Im April 2017 wurde über den Fund von vier neuen Riesenviren in der Kläranlage Klosterneuburg berichtet. Michael Wagner (Department für Mikrobiologie und Ökosystemforschung der Universität Wien), Holger Daims, Matthias Horn und Frederik Schulz (alle Uni Wien) publizierten über diese Klosneuviren (vorgeschlagene Unterfamilie der Mimiviridae) in Science.[46]
- „Orpheovirus“: Im Januar 2018 wurde von Julien Andreani und Kollegen über ein neues Riesenvirus Orpheovirus berichtet. Nach Vorschlag der Autoren soll „Orpheovirus“ in eine eigene Familie „Orpheoviridae“ gestellt werden, in enger Verwandtschaft zur vorgeschlagenen Familie Pithoviridae der Pithoviren.[47]


- „Meelsvirus“: Im September 2018 fanden Shinn und Bullard bei der nochmaligen Analyse von elektronenmikroskopischen Aufnahmen aus den 1980er Jahren ein Riesenvirus („Meelsvirus“), das den Pfeilwurm Adhesisagitta hispida infiziert, im Zellkern repliziert und dessen Virionen bei einer Länge von 1.25 μm umhüllt sind. Wegen fehlender Genomdaten ist eine bessere Einordnung bislang nicht möglich.[48][49]
- Waldbodenviren: Im November 2018 berichteten Frederik Schulz und Kollegen über die Entdeckung von 16 neuen Riesenviren per Metagenomanalyse von Waldbodenproben, die sich nur teilweise bekannten Gruppen zuordnen zu lassen scheinen. Für diese wurden vorläufige Namen vergeben, die meist auf ihre Herkunft hinweisen, unter anderem „Faunusvirus sp.“ (nicht zu verwechseln mit der offiziellen Gattung Faunusvirus der Familie Chaseviridae), „Gaeavirus“, „Homavirus“, „Barrevirus“, ‚Edafosvirus‘, „Hyperionvirus“, „Harvfovirus“, „Terrestrivirus“, „Dasosvirus“, „Satyrvirus“ (alle Mimiviridae), sowie „Sylvanvirus“,[50] „Solivirus“[51] und „Solumvirus“.[52] Möglicherweise ist dies nur die „Spitze eines Eisbergs“.[53][54]
- „Medusaviridae“: Ein Vorschlag für eine weitere Familie „Medusaviridae“ mit Gattung „Medusavirus“[6] findet sich beim CNRS (2018).[7][8][5][55] Diese Gruppe könnte basal von den NCLDV abzweigen und damit vielleicht einen zusätzlichen vierten Zweig bilden,[56] oder in der Nähe vom „Mollivirus“ angesiedelt sein.[5]
- „Yasminevirus“ etc.: Im März/April 2019 berichteten Clara Rolland und Kollegen in einer Zusammenfassung von Forschungsergebnissen über Riesenviren erstmals über weitere Kandidaten:[57] „Clandestinovirus“, „Fadolivirus“, „Sissivirus“, „Usurpativirus“, „Yasminevirus“[58] und „Misannotatedvirus“ (informell auch „misidentified virus“);[59] einem „mine drainage virus“;[59] sowie dem Virophagen Sissivirophage.
- „Choanovirus“: Im September 2019 berichteten David M. Needhal, Alexandra Z. Worden et al. über zwei Spezies (1 und 2) einer weiteren neuen Gattung „Choanovirus“ der erweiterten Mimiviren (jetzt Imitervirales). Der nächste Verwandte könnte das „Aureococcus anophagefferens virus“ („AaV“) sein.[60][61]
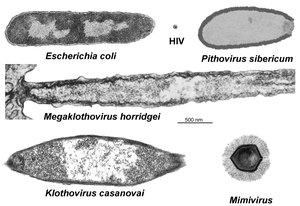
- „Klothoviridae“: Ebenfalls 2019 schlugen Roxane-Marie Barthélémy et al. eine Familie „Klothoviridae“ mit Typusspezies „Klothovirus casanovai“ und einer weiteren Spezies „Megaklothovirus horridgei“ vor, die mit 2,5–3,1 μm und 4 μm einen neuen Größenrekord darstellen würden. Als Wirte dienen wie bei „Meelsvirus“ Pfeilwürmer. Ähnlich wie bei den Arenaviridae fanden sich Ribosomen in den Viruspartikeln. Da derzeit noch keine Genomdaten vorliegen ist eine genauere Einordnung der Familie noch nicht möglich.[49]
.large.TMEs.jpg)
(D–F) Die Aufnahmen zeigen vergrößerte Zellkerne mit sich entwickelnden Viroplasmen.
(G–I) Die Morphologien der drei Viren umfassen einen Genom-Kern, umgeben von einem ikosaedrischen Nukleokapsid.

- „Mininucleoviridae“: Im Januar 2020 wurde von Subramaniam et al. eine weitere Familie „Mininucleoviridae“ von Riesenviren der NCLDV vorgeschlagen, deren Mitglieder Krebstiere (Crustacea) parasitieren. Zu den Mitgliedern dieser Familie gehören nach Vorschlag „Carcinus maenas virus 1“ (CmV1), „Dikerogammarus haemobaphes virus 1“ (DhV1) und „Panulirus argus virus 1“ (PaV1). Die Familie gehört offenbar zum Pitho-Irido-Marseille-Zweig der NCLDV (jetzt Ordnung Pimascovirales).[63]
- „Yaravirus“: Ebenfalls im Januar 2020 erschien ein Preprint von Paulo V. M. Boratto, Bernard La Scola, Jônatas S. Abrahão et al., in dem sie von einem DNA-Virus mit teilweise stark von den übrigen NCLDV abweichenden Genen berichteten, für das sie den Namen „Yaravirus brasiliensis“ vorschlagen.[11][12][64] Die Zugehörigkeit zu den NCLDV ist noch unklar, wegen der „double jelly roll“-Struktur wären diese jedoch nach ICTV den Bamfordvirae zuzuordnen.
- Im Oktober 2020 wurde eine Metagenomanalyse veröffentlicht, die zeigte, dass die Korallenbleiche bei Mo'orea, Franz. Polynesien, im Zusammenhang mit Viren steht. Obwohl eine eindeutige Zuordnung des rekonstruierten „assembled coral giant virus“ zu einem bestimmten Vertreter der NCLDV nicht möglich war, steht eine Zugehörigkeit zu dieser Gruppe außer Frage. Weitere Untersuchungen sind nötig.[65]
Systematik nach ICTV 2020

• Mimiviridae ist im erweiterten Sinn zu verstehen (Ordnung Imitervirales),
• die Asfar- und Poxviridae bilden eine Klade (Klasse Pokkesviricetes),
• die Ascovirdae sind paraphyletisch und bilden mit den Iridoviridae eine Klade („Irido-Ascoviridae“)
.svg)
Das Original wurde um die neuen ICTV-Bezeichnungen und den vorgeschlagenen Familiennamen ergänzt.
Das ICTV ist im März 2020 dem Vorschlag von Koonin et al. (2015 und 2019) einer inneren Systematik der NCLDV gefolgt, in der eine erweiterte Familie der Asfarviridae (jetzt Ordnung Asfuvirales) eine Schwestergruppe der Poxviridae (mit Ordnung Chitovirales) bilden. Von Koonin et al. (2015 und 2019) stammt die damit verträgliche Gliederung in drei Hauptgruppen (oder Zweige, englisch branches) wie folgt:[14][15][1]
- Phylum Nucleocytoviricota (Nucleocytoplasmic large DNA viruses (NCLDV), ursprünglicher Vorschlag „Nucleocytoplasmaviricota“,[2] ersetzt ältere Vorschläge als Ordnung „Megavirales“ s. l.)
- Klasse Megaviricetes
- Zweig 1:
- Ordnung Algavirales: die Familie Phycodnaviridae mit den Pandoraviren, „Mollivirus“, „Clandestinovirus“ und „Usurpativirus“,[57] aber ohne umgruppierte Vertreter wie die „OLPG“ (Organic Lake Phycodna group)
- Ordnung Imitervirales (veraltet „Megavirales“ s. s.[19]): die erweiterte Familie Mimiviridae s. l. syn. „Megaviridae“,[67] per Vorschlag mit „OLPG“ etc.
- Ordnung Algavirales: die Familie Phycodnaviridae mit den Pandoraviren, „Mollivirus“, „Clandestinovirus“ und „Usurpativirus“,[57] aber ohne umgruppierte Vertreter wie die „OLPG“ (
- Zweig 2: Ordnung Pimascovirales (Zusammenziehung aus Pitho, Irido, Marseille und Asco, früher auch MAPI-Superklade[19] oder PMI-Gruppe[28] genannt). Von oben nach unten immer basaler werdend gehören dazu:[63]
- die Familien Iridoviridae und Ascoviridae (derzeit vom ICTV noch getrennt aufgestellt, per Vorschlag eine gemeinsame Familie „Asco-Iridoviridae“),[47] ggf. mit Gattung Ichnovirus (bisher Polydnaviridae)
- Familie Marseilleviridae
- die vorgeschlagene Familie „Mininucleoviridae“[63]
- die vorgeschlagene Familie „Pithoviridae“ (mit „Cedratvirus“,[68][47] „Orpheoviridae“,[47] „Solumvirus“, „Solivirus“,[53] „Sissivirus“, „Misannotatedvirus“ (informell auch „misidentified virus“)[57][59] und „mine drainage virus“[59])
- Zweig 1:
- Klasse Pokkesviricetes
- Zweig 3:
- Ordnung Asfuvirales: (erweiterte) Familie Asfarviridae mit „Faustovirus“, „Pacmanvirus“,[69] „Kaumoebavirus“, evtl. Dinodnavirus,[42] etc.
- Ordnung Chitovirales: Familie Poxviridae (Pockenviren)
- Zweig 3:
- Klasse Megaviricetes
In dieses Schema wurden die Kandidaten nach Schulz et al. (2018), Rolland et al. (2019) und Subramaniam (2020) eingetragen, soweit sie nicht zu den erweiterten Mimiviridae (Ordnung Imitervirales) gehören. Die Kandidaten Dinodnavirus,[42] „Medusaviridae“, „Urceolovirus“ und „Yaravirus“ sind vom ICTV noch nicht berücksichtigt, was dem Vorschlag von Koonin et al. vom April 2019 entspricht.
Guglielmini et al. (2019) schlugen die Einteilung in zwei Superkladen folgendermaßen ein:[19][9]
- MAPI-Superklade (Zweig 2 von oben: Pimascovirales)
- PAM-Superklade (entspricht in etwa Zweig 1 und 3 (nur Asfuvirales))
Die Poxviridae (Pockenviren) sind in diesem Vorschlag unberücksichtigt.
Nach dem früheren Vorschlag von Schulz et al. (2018), Fig. 2,[53] stehen die Poxviridae dagegen basal in den NCLDV, die (erweiterten) Asfarviridae wurden im Zweig der Marseilleviridae verortet. Im Gegensatz zu Koonin et al. (2015 und 2019) erschienen die Riesenviren hier noch wie bei Guglielmini et al. monophyletisch. Das ICTV ist 2020 jedoch den Ergebnissen von Koonin, Yutin, Bäckström, Ettema et al. gefolgt, nachdem sich Riesenviren innerhalb der NCLDV mehrmals aus einfachen Vorstufen entwickelt haben:[14][28]
Nach Clara Rolland et al. (2019), Fig. 2[57] bilden die Imitervirales mit den Pokkesviricetes eine gemeinsame Klade, Schwestergruppen sind die Algavirales und Pimascovirales gleichauf. Die Megaviricetes scheinen – obgleich im März 2020 vom ICTV als Taxon offiziell bestätigt – keine Klade als Schwestergruppe der Pokkesviricetes zu bilden.
Nach Disa Bäckström et al. (2019), Fig. 1, scheinen – im Gegensatz zu Fig. 6 – die Pokkesviricetes ähnlich wie bei Schulz et al. (2018) keine gemeinsame Klade zu bilden, die Poxviridae (bzw. Chitovirales) stehen basal, erst danach zweigen die Asfarviridae (bzw. Asfuvirales) vom Rest der NCLDV ab.[28]
Weitere Stammbäume basierend auf Neighbor-joining und auf Maximum likelihood findet man u. a. in den folgenden Arbeiten:
- William H. Wilson et al. (2017), Fig. 2[70]
- Claire Bertelli et al. (2017), Fig. 4[68]
- Disa Bäckström (2018), Fig. 2[71]
- Disa Bäckström et al. (2019), Fig. 1 und 6[28]
- Sailen Barik (2018), Fig. 1[67]
Äußere Systematik
Die Organisation des Genoms und der DNA-Replikationsmechanismus legen eine phylogenetische Beziehung nahe zwischen den Rudiviren (Ordnung Ligamenvirales: Rudiviridae) und großen eukaryalen DNA-Viren (NCLDVs) wie dem Afrikanische Schweinepestvirus (African swine fever virus, Asfarviridae), Chloroviren (Chlorella virus, Phycodnaviridae) und Pockenviren (Orthopoxvirus, Poxviridae).[72]
Koonin et al. (2015, 2019) vermuten den Ursprung der NCLDV in den Tectiviridae, ikosaedrischen schwanzlosen ssDNA-Bakteriophagen, nach ICTV ebenfalls im Reich Bamfordvirae – im Unterschied zu den Herpesvirales (Herpesviren), bei denen der Ursprung bei den geschwänzten Caudovirales vermutet wird. Die Entwicklung verlief nach diesem Vorschlag über oder vermittels von Polintoviren (Polintons, auch Mavericks genannt: große DNA-Transposons, die virale Proteine kodieren, aber auch häufig in eukaryotischen Genomen vorkommen). Auch die Entwicklung von Adenoviren (Adenoviridae) und Bidnaviren (Bidnaviridae) sowie von Virophagen (der Familie Lavidaviridae) wurde, so die Vermutung, durch die Polintons initiiert.[14][15] Die im März 2020 vom ICTV getroffenen taxonomischen Einordnungen tragen dem Rechnung: sie ordnen
- die Herpesvirales und Caudovirales dem Reich Heunggongvirae im Bereich Duplodnaviria zu;
- die Tectiviridae, Adenoviridae (beide Klasse Tectiliviricetes) und Lavidaviridae (Klasse Maveriviricetes) dem Phylum Preplasmaviricota, einem Schwesterphylum der NCLDVs im Bereich Varidnaviria zu;
- lediglich die Bidnaviridae wurden per Vorschlag – ihrem Namen zum Trotz – dem Bereich Monodnaviria zugeordnet.[1]
Literatur
- Natalya Yutin, Yuri I. Wolf, Eugene V. Koonin: Origin of giant viruses from smaller DNA viruses not from a fourth domain of cellular life. In: Virology. 2014, doi:10.1016/j.virol.2014.06.032, PMC 4325995 (freier Volltext).
- N. Yutin, P. Colson, D. Raoult, E. V. Koonin: Mimiviridae: clusters of orthologous genes, reconstruction of gene repertoire evolution and proposed expansion of the giant virus family. In: Virol J. 10, 4. April 2013, S. 106, PMID 23557328
Weblinks
- Dagmar Röhrlich: Im Reich der Riesenviren. Deutschlandfunk – Wissenschaft im Brennpunkt, 27. Dezember 2015.
Anmerkungen
Einzelnachweise
- ↑ a b c d e f g ICTV: ICTV Master Species List 2019.v1, New MSL including all taxa updates since the 2018b release, March 2020 (MSL #35)
- ↑ a b c EV Koonin, VV Dolja, M Krupovic, A Varsani, YI Wolf, N Yutin, M Zerbini, JH Kuhn: Create a megataxonomic framework for DNA viruses encoding vertical jelly roll-type major capsid proteins filling all principal taxonomic ranks. ICTV Proposal 2019.003G, April–Juli 2019
- ↑ Jan Diesend, Janis Kruse, Monica Hagedorn, Christian Hammann: Amoebae, Giant Viruses, and Virophages Make Up a Complex, Multilayered Threesome. In: Frontiers in Cellular and Infection Microbiology, 7, Januar 2018, [doi:10.3389/fcimb.2017.00527], via ResearchGate, Fig. 1 („Megavirales“ im Sinn von NCLDV und NCLDV selbst werden in dieser Arbeit nicht ganz korrekt als ‚Familie‘ bezeichnet, gemeint ist ‚Gruppe‘).
- ↑ a b c d e Genki Yoshikawa, Romain Blanc-Mathieu, Chihong Song, Yoko Kayama, Tomohiro Mochizuki, Kazuyoshi Murata, Hiroyuki Ogata, Masaharu Takemura: Medusavirus, a novel large DNA virus discovered from hot spring water. In: Journal of Virology. 93, Nr. 8, 2019. doi:10.1128/JVI.02130-18. PMID 30728258. Abgerufen am 2. Juli 2019. jvi.asm.org (PDF)
- ↑ a b Daniel Lingenhöhl: Riesenvirus verwandelt Wirt zu Stein, auf: Spektrum.de vom 8. März 2019
- ↑ a b List of the main “giant” viruses known as of today (April, 18th, 2018) (PDF; 334 kB) Centre national de la recherche scientifique, Université Aix Marseille, 18. April 2018. Nach Masaharu Takemura et al.: Giant Viruses Isolated from Japanese Aquatic Environments, Tokyo University of Science, 3rd Ringberg Symposium on Giant Virus Biology (PDF) 19.–22. November 2017 (unveröffentlicht)
- ↑ a b List of the main “giant” viruses known as of today (March 2019). (PDF) Centre national de la recherche scientifique, Université Aix Marseille, März 2019.
- ↑ a b c Masaharu Takemura: Medusavirus Ancestor in a Proto-Eukaryotic Cell: Updating the Hypothesis for the Viral Origin of the Nucleus. In: Front. Microbiol., 11, S. 571831, 3. September 2020, doi:10.3389/fmicb.2020.571831
- ↑ a b Yetunde Adegboye, Mazen Atia, Anna M. Sigmund, Anne Krüger, Carsten Kirschning et al.: Abundance and Immunogenicity of Two Giant Viruses Namely Pithovirus lacustris and Urceolovirus corneum; researchgate.net (PDF). Conference Paper, 14th International Conference on Microbial Interactions & Microbial Ecology, Vienna (Austria), August 2019
- ↑ a b Paulo V. M. Boratto, Graziele P. Oliveira, Talita B. Machado, Ana Cláudia S. P. Andrade, Jean-Pierre Baudoin, Thomas Klose, Frederik Schulz, Saïd Azza, Philippe Decloquement, Eric Chabrière, Philippe Colson, Anthony Levasseur, Bernard La Scola, Jônatas S. Abrahão: A mysterious 80 nm amoeba virus with a near-complete “ORFan genome” challenges the classification of DNA viruses, auf: bioRxiv, 28. Januar 2020, bioRxiv: 10.1101/2020.01.28.923185v1 (Preprint-Volltext) doi:10.1101/2020.01.28.923185
- ↑ a b Paulo V. M. Boratto, Graziele P. Oliveira, Talita B. Machado, Ana Cláudia S. P. Andrade, Jean-Pierre Baudoin, Thomas Klose, Frederik Schulz, Saïd Azza, Philippe Decloquement, Eric Chabrière, Philippe Colson, Anthony Levasseur, Bernard La Scola, Jônatas S. Abrahão; James L. Van Etten (Hrsg.): Yaravirus: A novel 80-nm virus infecting Acanthamoeba castellanii. In: PNAS, Band 117 Nr. 28, 14. Juli 2020, S. 16579–16586, online seit 29. Juni 2020 (IF 9.412), X-mol, doi:10.1073/pnas.2001637117
- ↑ Lakshminarayan M. Iyer, L. Aravind, Eugene V. Koonin: Common Origin of Four Diverse Families of Large Eukaryotic DNA Viruses. In: Journal of Virology. 75, 2001, S. 11720–11734, doi:10.1128/JVI.75.23.11720-11734.2001, PMC 114758 (freier Volltext).
- ↑ a b c d e f g Eugene V. Koonin, Natalya Yutin: Evolution of the Large Nucleocytoplasmatic DNA Viruses of Eukaryotes and Convergent Origins of Viral Gigantism. In: Advances in Virus research, Band 103, AP 21. Januar 2019, doi:10.1016/bs.aivir.2018.09.002, S. 167–202. Die Klosneuviren sind teilweise als Klosneviren fehlgeschrieben.
- ↑ a b c Eugene V. Koonin, Valerian V. Dolja, Mart Krupovic: Origins and evolution of viruses of eukaryotes: The ultimate modularity. In: Virology, Mai 2015, S. 479–480. 2–25, Epub 12. März 2015, PMC 5898234 (freier Volltext), PMID 25771806
- ↑
Natalya Yutin, Yuri I. Wolf, Eugene V. Koonin: Origin of giant viruses from smaller DNA viruses not from a fourth domain of cellular life. In: Virology, Oktober 2014, doi:10.1016/j.virol.2014.06.032, PMID 25042053: „… we refer to this major group of viruses as ‘Megavirales’ to signal our support of this amendment to virus taxonomy …“, einem Vorschlag aus der Arbeitsgruppe um Didier Raoult folgend, siehe P. Colson, X. de Lamballerie, G. Fournous, D. Raoult: Reclassification of giant viruses composing a fourth domain of life in the new order Megavirales. In: Intervirology, 55, 2012, S. 321–332, doi:10.1159/000336562, PMID 22508375.
- ↑ ICTV Executive Committee: The new scope of virus taxonomy: partitioning the virosphere into 15 hierarchical ranks. In: Nature Microbiology, 5, S. 668–674, 27. April 2020, doi:10.1038/s41564-020-0709-x, researchgate.net Stand: Januar 2020
- ↑ Nadja Podbregar: Ein Stammbaum für die Virosphäre. scinexx.de, 29. April 2020. Stand: Januar 2020
- ↑ a b c d Julien Guglielmini, Anthony C. Woo, Mart Krupovic, Patrick Forterre, Morgan Gaia: Diversification of giant and large eukaryotic dsDNnA viruses predated the origin of modern eukaryotes. In: PNAS, Band 116, Nr. 39, 10./24. September 2019, S. 19585–19592, doi:10.1073/pnas.1912006116, PMID 31506349, Fig. 2. Dazu:
Julien Guglielmini, Anthony Woo, Mart Krupovic, Patrick Forterre, Morgan Gaia: Diversification of giant and large eukaryotic dsDNA viruses predated the origin of modern eukaryotes, auf: bioRxiv vom 29. Oktober 2018, bioRxiv: 10.1101/455816v1 (Preprint-Volltext) (PrePrint) - ↑
N. Yutin, Y. I. Wolf, E. V. Koonin: Origin of giant viruses from smaller DNA viruses not from a fourth domain of cellular life. In: Virology, 2014: „all giant viruses contain a set of core genes that define … the proposed order Megavirales“.
- ↑ a b James L. Van Etten, Leslie C. Lane, David D. Dunigan: DNA Viruses: The Really Big Ones (Giruses). Annu. Rev. Microbiol. 64, 2010, S. 83–99, doi:10.1146/annurev.micro.112408.134338 (Volltext frei zum persönlichen Gebrauch (PDF; 402 kB) ).
- ↑ Viruses don’t have a metabolism; but some have the building blocks for one (en). In: phys.org. Abgerufen am 15. Mai 2020.
- ↑ Mohammad Moniruzzaman, Carolina A. Martinez-Gutierrez, Alaina R. Weinheimer, Frank O. Aylward: Dynamic genome evolution and complex virocell metabolism of globally-distributed giant viruses. In: Nature Communications. 11, Nr. 1, 6. April 2020, ISSN 2041-1723, S. 1710. bibcode:2020NatCo..11.1710M. doi:10.1038/s41467-020-15507-2. PMID 32249765. PMC 7136201 (freier Volltext).
- ↑ Natalya Yutin, Philippe Colson, Didier Raoult, Eugene V. Koonin: Mimiviridae: clusters of orthologous genes, reconstruction of gene repertoire evolution and proposed expansion of the giant virus family. In: Virol J., 10, 2013, S. 106, doi:10.1186/1743-422X-10-106. PMID 23557328.
- ↑ a b Anthony Levasseur, Meriem Bekliz, Eric Chabrière, Pierre Pontarotti, Bernard La Scola und Didier Raoult: MIMIVIRE is a defence system in mimivirus that confers resistance to virophage. In: Nature. 2016, doi:10.1038/nature17146 (freier Volltext).
- ↑ d. h. intrazellulärer Symbiont oder Parasit
- ↑ Natalya Yutin, Yuri I. Wolf, Eugene V. Koonin: Origin of giant viruses from smaller DNA viruses not from a fourth domain of cellular life. In: Virology, 2014, Fig. 1 bis 4.
- ↑ a b c d e Disa Bäckström, Natalya Yutin, Steffen L. Jørgensen, Jennah Dharamshi, Felix Homa, Katarzyna Zaremba-Niedwiedzka, Anja Spang, Yuri I. Wolf, Eugene V. Koonin, Thijs J. G. Ettema; Richard P. Novick (Hrsg.): Virus Genomes from Deep Sea Sediments Expand the Ocean Megavirome and Support Independent Origins of Viral Gigantism, in: mBio Vol. 10, Nr. 2, März–April 2019, S. e02497-18, mbio.asm.org (PDF) doi:10.1128/mBio.02497-18, PMC 6401483 (freier Volltext), PMID 30837339, ResearchGate. Anm.: In Fig. 1 muss es Megamimivirinae (oder Megavirinae) statt Mimiviridae heißen. Diese Unterfamilie bildet zusammen mit den Klosneuvirinae und den Cafeteriaviren die Familie Mimiviridae s. s.
- ↑ Traci Watson: The trickster microbes that are shaking up the tree of life. In: Nature, 14. Mai 2019 (englisch), Trickser-Bakterien schütteln den Stammbaum des Lebens. Spektrum.de, 20. Juni 2019 (deutsch) – die Bezeichnung ‚Bakterien‘ ist nicht ganz korrekt, bei den betrachteten Mikroben handelt es sich um Archaeen oder (nach Ansicht mancher Forscher) jedenfalls um von den Bakterien verschiedene Proto-Eukaryonten.
- ↑ Patrick L. Scheid: Free-Living Amoebae and Their Multiple Impacts on Environmental Health. In: Reference Module in Earth Systems and Environmental Sciences, 27. Februar 2018, doi:10.1016/B978-0-12-409548-9.10969-8, hier: Text im Anschluss an Fig. 8 (rechte Spalte)
- ↑ Florian Maumus, Aline Epert, Fabien Nogué, Guillaume Blanc: Plant genomes enclose footprints of past infections by giant virus relatives, Nature Communications Band 5, Nr. 4268, 27. Juni 2014, doi:10.1038/ncomms5268
- ↑ NCBI: Cotesia vestalis bracovirus (species) – Vorschlag.
- ↑ Paul B. Talbert, Karim-Jean Armache, Steven Henikoff: Viral histones: pickpocket’s prize or primordial progenitor? In: BMC Epigenetics & Chromatin, Band 15, Nr. 21, 28. Mai 2022; doi:10.1186/s13072-022-00454-7, PMID 35624484, PMC 9145170 (freier Volltext).
- ↑ Takashi Yamada: Giant viruses in the environment: Their origins and evolution. In: Curr Opin Virol., Juli 2011;1(1), S. 58–62, doi:10.1016/j.coviro.2011.05.008, PMID 22440568
- ↑ N. Yutin, Y. I. Wolf, E. V. Koonin: Origin of giant viruses from smaller DNA viruses not from a fourth domain of cellular life. In: Virology, 2014, Fig. 5 und 6.
- ↑ K. Stasiak, M. V. Demattei, B. A. Federici, Y. Bigot: Phylogenetic position of the Diadromus pulchellus ascovirus DNA polymerase among viruses with large double-stranded DNA genomes. In: The Journal of General Virology. Band 81, Pt 12, Dezember 2000, S. 3059–72, doi:10.1099/0022-1317-81-12-3059, PMID 11086137.
- ↑ K. Stasiak, S. Renault, M. V. Demattei, Y. Bigot, B. A. Federici: Evidence for the evolution of ascoviruses from iridoviruses. In: The Journal of General Virology. Band 84, Pt 11, November 2003, S. 2999–3009, doi:10.1099/vir.0.19290-0, PMID 14573805.
- ↑ a b B. A. Federici, D. K. Bideshi, Y. Tan, T. Spears, Y. Bigot: Ascoviruses: superb manipulators of apoptosis for viral replication and transmission. In: Current Topics in Microbiology and Immunology. Band 328, 2009, ISBN 978-3-540-68617-0, S. 171–96, doi:10.1007/978-3-540-68618-7_5, PMID 19216438.
- ↑ B. Piégu, S. Asgari, D. Bideshi, B. A. Federici, Y. Bigot: Evolutionary relationships of iridoviruses and divergence of ascoviruses from invertebrate iridoviruses in the superfamily Megavirales. In: Molecular Phylogenetics and Evolution. Band 84, März 2015, S. 44–52, doi:10.1016/j.ympev.2014.12.013, PMID 25562178.
- ↑ dsDNA Viruses > Ascoviridae, auf: ICTV online, Dezember 2016 (hier: Fig. 2)
- ↑ Fenner’s Veterinary Virology (Fifth Edition): Chapter 8 – Asfarviridae and Iridoviridae, online 4. November 2016, S. 175–188, doi:10.1016/B978-0-12-800946-8.00008-8 (hier: Fig. 1)
- ↑ a b c Hiroyuki Ogata, Kensuke Toyoda, Yuji Tomaru, Natsuko Nakayama, Yoko Shirai, Jean-Michel Claverie, Keizo Nagasaki: Remarkable sequence similarity between the dinoflagellate-infecting marine girus and the terrestrial pathogen African swine fever virus. In: Virol J., 6, 2009, doi:10.1186/1743-422X-6-178, S. 178.
- ↑ Urceolovirus corneum (species). NCBI
- ↑ SAMN06480674 – U. corneum nucleotide, auf: BioSamples, European Bioinformatics Institute (GB)
- ↑ Urceolovirus corneum (species), auf: DDBJ DNA Data bank of Japan
- ↑ Forscher rätseln über Riesenviren aus Kläranlage orf.at, 6. April 2017, abgerufen 6. April 2017.
- ↑ a b c d Julien Andreani, Jacques Y. B. Khalil, Emeline Baptiste, Issam Hasni, Caroline Michelle, Didier Raoult, Anthony Levasseur, Bernard La Scola: Orpheovirus IHUMI-LCC2: A New Virus among the Giant Viruses. In: Frontiers in Microbiology. Band 8, 22. Januar 2018, ISSN 1664-302X, doi:10.3389/fmicb.2017.02643 (frontiersin.org [PDF]).
- ↑ a b c G. L. Shinn, B. L. Bullard; Carmen San Martin (Hrsg.): Ultrastructure of Meelsvirus: A nuclear virus of arrow worms (phylum Chaetognatha) producing giant “tailed” virions. In: PLOS ONE. 13, Nr. 9, 19. September 2018, S. e0203282. bibcode:2018PLoSO..1303282S. doi:10.1371/journal.pone.0203282. PMID 30231047. PMC 6145532 (freier Volltext).
- ↑ a b c Roxane-Marie Barthélémy, Eric Faure, Taichiro Goto: Serendipitous Discovery in a Marine Invertebrate (Phylum Chaetognatha) of the Longest Giant Viruses Reported till Date. In: Biology, 2019, Abstract, hilarispublisher.com (PDF; 1,3 MB)
- ↑ Sylvanvirus (species). NCBI
- ↑ Solivirus (species). NCBI
- ↑ Solumvirus (species). NCBI
- ↑ a b c Frederik Schulz, Lauren Alteio, Danielle Goudeau, Elizabeth M. Ryan, Feiqiao B. Yu, Rex R. Malmstrom, Jeffrey Blanchard, Tanja Woyke: Hidden diversity of soil giant viruses. In: Nature Communications, volume 9, Article number: 4881, 19. November 2018; doi:10.1038/s41467-018-07335-2.
- ↑ Jan Osterkamp: Virologie: Riesenviren sind weiter verbreitet als gedacht. Spektrum.de, 20. November 2018.
- ↑ The giant Medusavirus turns defenceless cells to 'stone'. In: Nature. 566, Nr. 7745, 2019, S. 429. bibcode:2019Natur.566R.429.. doi:10.1038/d41586-019-00591-2. Abgerufen am 2. Juli 2019.
- ↑ New giant virus may help scientists better understand the emergence of complex life – Large DNA virus that helps scientists understand the origins of DNA replication and the evolution of complex life. In: EurekAlert!, Tokyo University of Science, 30. April 2019. Abgerufen am 2. Juli 2019.
- ↑ a b c d Clara Rolland, Julien Andreani, Amina Cherif Louazani, Sarah Aherfi, Rania Francis, Rodrigo Rodrigues, Ludmila Santos Silva, Dehia Sahmi, Said Mougari, Nisrine Chelkha, Meriem Bekliz, Lorena Silva, Felipe Assis, Fábio Dornas, Jacques Yaacoub Bou Khalil, Isabelle Pagnier, Christelle Desnues, Anthony Levasseur, Philippe Colson, Jônatas Abrahão, Bernard La Scola: Discovery and Further Studies on Giant Viruses at the IHU Mediterranee Infection That Modified the Perception of the Virosphere. In: Viruses, 11(4), März/April 2019, pii: E312, doi:10.3390/v11040312, PMC 6520786 (freier Volltext), PMID 30935049.
- ↑ Leena Hussein Bajrai, Saïd Mougari, Julien Andreani, Emeline Baptiste, Jeremy Delerce, Didier Raoult, Esam Ibraheem Azhar, Bernard La Scola, Anthony Levasseur; Joanna L. Shisler (Hrsg.): Isolation of Yasminevirus, the First Member of Klosneuvirinae Isolated in Coculture with Vermamoeba vermiformis, Demonstrates an Extended Arsenal of Translational Apparatus Components. In: JVirol, 94(1), Herbst 2019, doi:10.1128/JVI.01534-19
- ↑ a b c d Julien Andreani, Jonathan Verneau, Didier Raoult, Anthony Levasseurn Bernard La Scola: Deciphering viral presences: two novel partial giant viruses detected in marine metagenome and in a mine drainage metagenome, in: Virology Journal, Band 15, Nr. 66, 10. April 2018, doi:10.1186/s12985-018-0976-9
- ↑ David M. Needham, Susumu Yoshizawa, Toshiaki Hosaka, Camille Poirier, Chang Jae Choi, Elisabeth Hehenberger, Nicholas A. T. Irwin, Susanne Wilken, Cheuk-Man Yung, Charles Bachy, Rika Kurihara, Yu Nakajima, Keiichi Kojima, Tomomi Kimura-Someya, Guy Leonard, Rex R. Malmstrom, Daniel R. Mende, Daniel K. Olson, Yuki Sudo, Sebastian Sudek, Thomas A. Richards, Edward F. DeLong, Patrick J. Keeling, Alyson E. Santoro, Mikako Shirouzu, Wataru Iwasaki, Alexandra Z. Worden: A distinct lineage of giant viruses brings a rhodopsin photosystem to unicellular marine predators. In: PNAS, 23. September 2019, doi:10.1073/pnas.1907517116, ISSN 0027-8424, pnas.org (PDF) inklusive Supplement 1 (xlsx)
- ↑ Mohammad Moniruzzaman, Alaina R. Weinheimer, Carolina A. Martinez-Gutierrez, Frank O. Aylward: Widespread endogenization of giant viruses shapes genomes of green algae. In: nature, 18. November 2020, doi:10.1038/s41586-020-2924-2, dazu:
Kendall Daniels: Lurking in genomic shadows: How giant viruses fuel the evolution of algae. In: vtnews, SciTechDaily. Virginia Tech, 18. November 2020 - ↑ Ökologie und Biodiversität des Kulkwitzer Sees, auf: Landessportbund Sachsen
- ↑ a b c d e f Kuttichantran Subramaniam, Donald C. Behringer, Jamie Bojko, Natalya Yutin, Abigail S. Clark, Kelly S. Bateman, Ronny van Aerle, David Bass, Rose C. Kerr, Eugene V. Koonin, Grant D. Stentiford, Thomas B. Waltzek; Vincent R. Racaniello (Hrsg.): A New Family of DNA Viruses Causing Disease in Crustaceans from Diverse Aquatic Biomes.. In: mBio. 11, Nr. 1, 14. Januar 2020. doi:10.1128/mBio.02938-19. PMID 31937645. PMC 6960288 (freier Volltext).
- ↑ Peter Dockrill: Scientists Discover Mysterious Virus in Brazil With No Known Genes They Can Identify, auf: sciencealert vom 10. Februar 2020
- ↑ Adriana Messyasz, Stephanie M. Rosales, Ryan S. Mueller, Teresa Sawyer, Adrienne M. S. Correa, Andrew R. Thurber, Rebecca Vega Thurber: Coral Bleaching Phenotypes Associated With Differential Abundances of Nucleocytoplasmic Large DNA Viruses. In: Frontiers in Marine Science, Band 6, Coral Reef Research, 6. Oktober 2020, S. 789, ISSN 2296-7745, doi:10.3389/fmars.2020.555474, frontiersin.org (PDF) Supplement, dazu:
- Steve Lundeberg: Oregon State University scientists shed new light on viruses’ role in coral bleaching. Oregon State University, Pressemitteilung, 12. Oktober 2020
- Scientists shed new light on viruses’ role in coral bleaching. EurekAlert!, 14. Oktober 2020
- ↑ Sudhir Kumar, Glen Stecher, Koichiro Tamura: MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. In: Molecular Biology and Evolution. 33, Nr. 7, 2016, S. 1870–1874. doi:10.1093/molbev/msw054. PMID 27004904.
- ↑ a b Sailen Barik: A Family of Novel Cyclophilins, Conserved in the Mimivirus Genus of the Giant DNA Viruses. In: Computational and Structural Biotechnology Journal, Band 16, Juli 2018, S. 231–236, doi:10.1016/j.csbj.2018.07.001
- ↑ a b Claire Bertelli, Linda Mueller, Vincent Thomas, Trestan Pillonel, Nicolas Jacquier, Gilbert Greub: Cedratvirus lausannensis – digging into Pithoviridae diversity. In: Environmental Microbiology, 2017, 19(10), S. 4022–4034, doi:10.1111/1462-2920.13813
- ↑ Pierre-Philippe Dechant: Recent developments in mathematical virology (PDF; 24 MB) ICERM, York St. John University, 15. November 2018
- ↑ William H. Wilson, Ilana C. Gilg, Mohammad Moniruzzaman, Erin K. Field, Sergey Koren, Gary R. LeCleir, Joaquín Martínez Martínez, Nicole J. Poulton, Brandon K. Swan, Ramunas Stepanauskas, Steven W. Wilhelm: Genomic exploration of individual giant ocean viruses. In: ISME Journal, 11(8), August 2017, S. 1736–1745, doi:10.1038/ismej.2017.61, PMC 5520044 (freier Volltext), PMID 28498373
- ↑ Disa Bäckström: Exploring the diversity and evolution of giant viruses in deep sea sediments using genome-resolved metagenomics. (PDF) Semantic Scholar, 2018
- ↑ D. Prangishvili, R. A. Garrett: Exceptionally diverse morphotypes and genomes of crenarchaeal hyperthermophilic viruses. In: Biochemical Society Transactions. Band 32, Pt 2, April 2004, S. 204–8, doi:10.1042/BST0320204, PMID 15046572.

{kind=link}
{kind=link}
{kind=link}
{kind=link}